|
|
Post by Admin on Oct 25, 2019 19:08:42 GMT
What does it mean to be “cute”? Beauty standards and perceptions of attractiveness vary across cultures, but one thing is clear: being called “cute” is something entirely different. Is there a scientific explanation of why our brains perceive some people as “cute” and others as “beautiful,” “hot,” and so on? We’re about to answer all of them with the help of science! TIMESTAMPS: Why we find babies so cute 1:13 What the EDAR gene is 2:28 What sexual dimorphism is 3:47 The gene in question is known as EDAR. Native Americans and Asians carry a version of the gene that is linked to thicker hair shafts, more sweat glands, and shovel-shaped incisors. A variant of this gene—V370A—arose about 30,000 years ago or so in China when the climate was hotter and more humid, which prompted researchers to speculate initially that it was advantageous to have more sweat glands in that environment. But the gene variant swept through the ancestors of Asians and Native Americans about 20,000 years ago, when the climate where they lived in Asia and Beringia (the now-submerged land between Asia and Alaska) was colder and dryer. So the actual cause of the gene’s spread has been unknown.  The new study reveals that the variant was so beneficial it spread to everyone in the Americas. When researchers led by biological anthropologist Leslea Hlusko of the University of California, Berkeley, examined data on the teeth of more than 5000 people from 54 archaeological sites in Europe, Asia, and North and South America, they found shoveled teeth—and hence, the gene variant that causes them—in about 40% of the individuals in Asia and all of the 3183 fossils of Native Americans they examined (who all lived before European colonization). This suggests that some members of the first group to arrive in Beringia probably carried the gene, which arose in Asia. Then it quickly swept through the rest of the small isolated population of people who settled there between 28,000 and 18,000 years ago.  Living at such a high latitude puts nursing infants at risk of not getting enough sunlight in winter to synthesize vitamin D in their skin. Unlike adults, nursing infants can’t eat marine foods and organ meat rich in vitamin D to compensate. Vitamin D deficiency can trigger serious problems with bone development, such as rickets. It also interferes with the many ways fat insulates and fuels our bodies, as well as how the immune system wards off disease. The EDAR variant led to the development of more elaborate branching of milk ducts in studies of mice. Hlusko and her colleagues hypothesize that those extra branches cause mothers to produce more milk or deliver more nutrients in their milk. If so, children of mothers with the EDAR variant would have been more likely to survive, thus spreading the variant throughout the population, the team proposes today in the Proceedings of the National Academy of Sciences.  The study shows how natural selection can work rapidly when humans move into extreme environments, such as the Arctic, exerting strong selection on genes critical for development and metabolism, says biological anthropologist William Leonard of Northwestern University in Evanston, Illinois, who was not involved with this work. Follow-up studies need to be done to explore just how an increase in breast ducts might deliver more nutrients. One possibility, Hlusko says, is that the EDAR gene variant works with fatty acid genes to deliver more fats in breast milk. Researchers need to see whether there is a connection between the genes, says geneticist Tábita Hünemeier of the University of São Paulo in Brazil, who found selection for fatty acid genes in Native Americans and is not involved with the work. Hlukso is now collaborating with others to explore how the EDAR gene variant affects breast development and density. “This is not just a Native American story,” Hlusko says. “Everyone with shovel-shaped incisors has this gene that may compensate for vitamin D deficiency.” |
|
|
|
Post by Admin on Oct 25, 2019 22:51:23 GMT
Because of the ubiquitous adaptability of our material culture, some human populations have occupied extreme environments that intensified selection on existing genomic variation. By 32,000 years ago, people were living in Arctic Beringia, and during the Last Glacial Maximum (LGM; 28,000–18,000 y ago), they likely persisted in the Beringian refugium. Such high latitudes provide only very low levels of UV radiation, and can thereby lead to dangerously low levels of biosynthesized vitamin D. The physiological effects of vitamin D deficiency range from reduced dietary absorption of calcium to a compromised immune system and modified adipose tissue function. The ectodysplasin A receptor (EDAR) gene has a range of pleiotropic effects, including sweat gland density, incisor shoveling, and mammary gland ductal branching. The frequency of the human-specific EDAR V370A allele appears to be uniquely elevated in North and East Asian and New World populations due to a bout of positive selection likely to have occurred circa 20,000 y ago. The dental pleiotropic effects of this allele suggest an even higher occurrence among indigenous people in the Western Hemisphere before European colonization. We hypothesize that selection on EDAR V370A occurred in the Beringian refugium because it increases mammary ductal branching, and thereby may amplify the transfer of critical nutrients in vitamin D-deficient conditions to infants via mothers’ milk. This hypothesized selective context for EDAR V370A was likely intertwined with selection on the fatty acid desaturase (FADS) gene cluster because it is known to modulate lipid profiles transmitted to milk from a vitamin D-rich diet high in omega-3 fatty acids.  Fig. 1. Geography of Beringia and levels of UV radiation. (A) Map of Beringia today. Cross-hatching indicates the region in which levels of UVMED (defined as the amount of UV radiation that will produce minimal erythema) that reach the Earth’s surface are too low to promote cutaneous synthesis of vitamin D in humans on a year-by-year basis, requiring dietary supplementation (modified from ref. 84; projected to show an equal distance map of Beringia). The black and white region marks the Arctic Circle, for which there are no Total Ozone Mapping Satellite Data version 7. Other data show that this region has even less UV-B exposure, as would be expected from the increased latitude. The areas below the Arctic Circle in white and light blue are shallow seas as discerned from modern bathymetry using Etopo2 data, indicating land that would have been exposed during the LGM. (B) Map of Beringia during the LGM showing the exposure of land at 117 m below current sea level and the reconstructed terrestrial environments. The shrub tundra is the only area biologically productive enough to support a human population of the size estimated by molecular data. This population was genetically isolated for ∼2,500–9,000 y during the LGM because of the ice to the east and extensive mesic tundra to the west. From Thomas Jefferson’s archaeological excavations (1) to modern genomics (2⇓–4), scientists have been fascinated by the first migration of humans into the Americas. A myriad of new evidence reveals that the earliest people in the Western Hemisphere dispersed from a population that lived in genetic isolation for thousands of years on the exposed Beringian platform in the Arctic during the Last Glacial Maximum [LGM; 28,000–18,000 y ago (2⇓⇓⇓⇓–7)]. The Arctic is an extreme environment because of the very low UV radiation (UV) reaching the earth’s surface at such high latitude. UV is essential to almost all life forms because it catalyzes biochemical processes, especially the synthesis of vitamin D (8).  Extreme environments can impact genetic variation and provide opportunities to elucidate relationships between genotype and phenotype (9). The classic human example is the range of our physiological adaptations to the hypoxic conditions of high altitude, such as are experienced in the highlands of Ethiopia, the Tibetan Plateau, and the Andean Mountains (10⇓–12). Evidence shows that the populations which have long-occupied these high-elevation regions have an increased frequency of red blood cell polymorphisms that likely underlie these physiologies (10⇓–12). Here, we investigate whether the population occupying Beringia during the LGM represents another example of human adaptation to an extreme environment, this time adapting to very low UV exposure (Fig. 1). There are two lines of genetic evidence for this: variation in the fatty acid desaturase (FADS) gene cluster that modulates the manufacture of polyunsaturated fatty acids and variation in the ectodysplasin A receptor (EDAR) gene that influences ectodermally derived structures, such as teeth, hair, and mammary gland ductal branching. A study on selection on the FADS gene cluster in the ancestral population of Native Americans has been published previously (13), but, here, we shift the emphasis from phenotypic effects on older adults to focus on those that influence fertility via breast milk. We then present evidence that EDAR may have undergone an episode of selection in the same population, likely due to its influence on mammary ductal branching. We hypothesize that the genetically isolated population which occupied Beringia during the LGM experienced selection for an increase in vitamin D in breast milk in response to the low UV environment. Traces of this previously intense selection appear to still be present in the genetic variation of Native American and North and East Asian populations today.  Origins of 370A (A) Haplotype distribution of the genomic region surrounding V370A , based on 24 SNPs covering $ 139 kb. The six most common haplotypes are shown, and the remaining low-frequency haplotypes grouped as ‘‘Other.’’ The chimpanzee allele was assumed to be ancestral. Derived alleles are in dark blue, except for the 370A variant which is red. |
|
|
|
Post by Admin on Oct 26, 2019 18:27:00 GMT
The FADS Gene Cluster Allelic variation in the FADS gene complex corresponds strongly to geographic ancestry (14, 15), and is the most pronounced allele frequency difference between the Greenlandic Inuit and other human populations (16). Because modern Western cultures place much attention on the cardiovascular disease risks associated with diets rich in omega-6 versus omega-3 long-chain polyunsaturated fatty acids (LC-PUFAs) (17, 18), the high frequencies of these alleles in the Inuit have been primarily interpreted as an adaptation to their traditionally meat-rich (and omega-3–rich) diet (16, 19). However, and surprisingly, 95% of native Central and South Americans also show evidence of selection on the same FADS polymorphisms, yet these cultural groups do not traditionally consume diets nearly as rich in omega-3 fatty acids as do the Inuit, suggesting that their common ancestral population is characterized by one which did (13). From an evolutionary perspective, the possibility that the selective force favoring these FADS alleles was a healthier ratio of omega-3 to omega-6 fatty acids in adults would indicate that the effect must have been very strong, as selective pressure is relatively low on genetic variation that influences later and post-reproductive years of life (20). In contrast, even very small positive effects on fertility and childhood survival have far greater selective power (20). Considering this, the selective benefits of these FADS polymorphisms are more likely connected to their significant role in modulating the relative levels of omega-3 and omega-6 fatty acids during gestation and postnatal growth of infants. To elaborate, the fatty acids arachidonic acid, docosahexaenoic acid (DHA), and eicosapentaenoic acid (EPA) are essential to cognitive and visual development (21). Maternal polymorphisms in the FADS gene cluster have strong effects on the levels of EPA and DHA in breast milk [in humans (22⇓⇓⇓⇓–27) and in other mammals (27, 28)]. This suggests that lipid levels in a mother’s diet are optimized by specific polymorphisms in her FADS gene cluster during milk synthesis (Fig. 2A). These polymorphisms are therefore a potentially strong target of selection because fetuses and infants have a very limited ability to synthesize LC-PUFAs on their own and are dependent on their mother’s genotype to modulate their relative proportions (22⇓⇓⇓⇓–27). We propose that the phenotypic effects of FADS polymorphisms on the LC-PUFA content of breast milk was the primary target of selection in past human populations that consumed diets with compromised proportions of omega-3 and omega-6 fatty acids, such as is seen in the traditional diets of people living in the Arctic.  Fig. 2. Overview of breast anatomy, development, milk production, and histology. (A) Genetic influences on milk production summarized from the main text. The rodent, human, and cow figures indicate the systems from which the results are known. (B) Three stages of mammary development alongside the genetic mutations known to increase ductal branching. Hormones induce the pubertal and gestation/lactation stages (denoted in pink). In vitamin D-deficient adipose tissue, ductal branching increases during these two stages of development. The embryonic stage (in green) is not hormonally induced; branching density is influenced by the ectodysplasin pathway (108) and, specifically, increased by EDAR V370A. (C) Cross-section of a ductal lumen showing the bilayer of luminal and myoepithelial cells. (D) Schematic of a desmosome, one of the main adhesive structures between mammary luminal cells. The orange lines represent desmosomal cadherins in the extracellular space. The gray lines in the purple area represent the intracellular filaments. (E) Close-up view of the desmocolin proteins that comprise the desmo-adhesome. An increase in the activity of the ectodyplasin pathway alters the relative proportions of Dsc2 and Dsc3, leading to a reduction in the adhesive strength. EDAR. The human EDAR V370A variant encodes a change in amino acid sequence in the highly conserved death domain of EDAR (29⇓⇓–32). EDAR functions as a protein receptor on a cell’s surface that activates the transcription factor NF-ĸB in the ectodysplasin pathway (29). Comparative studies show this pathway is functionally conserved across virtually all vertebrates, playing essential roles in the development of ectodermal structures from bird feathers to fish scales (33, 34). EDAR is one of four genes implicated in hypohidrotic ectodermal dysplasia (HED), a set of ∼150 syndromes characterized by mild to severe defects in ectodermally derived structures, such as hair, teeth, breasts, and sweat glands (33, 35⇓–37). In contrast to HED, the EDAR V370A allele has the opposite effect. Genome-wide association studies (GWASs) of multiple Asian populations show that EDAR V370A is correlated with hair shaft caliber (38, 39), earlobe and chin shape (40), and a suite of morphological variants on teeth (40⇓⇓–43). Knock-in mouse studies reveal that the V370A allele leads to a twofold increase in NF-ĸB activation (30, 31, 44). Just like humans, the EDAR V370A mouse has thicker hair shafts, an increase in the branching density of mammary gland ducts, and (inconsistently) an increase in the number of eccrine glands on the footpads (30, 31). Several studies conclude that EDAR V370A experienced a bout of intense positive selection ∼20,000 y ago in northern China (29, 31, 45). Previous interpretations as to why this selection occurred focused on the associated increase in eccrine sweat gland density on the fingertips, concluding that the selection was for improved thermoregulatory  during a warm spell during the LGM (31) or that the increase in sebaceous glands would offer protection from the cold, dry air of the LGM (30). These interpretations rely primarily on the current distribution of the allele in living populations (Fig. 3A), which shows high frequencies of EDAR V370A in North and East Asian and Native American populations but virtual absence in other populations around the world (29⇓–31, 45).  Fig. 3. Relationship between EDAR V370A and degree of incisor shoveling. (A) Current world-wide allele frequencies of EDAR V370A (in yellow) and other EDAR haplotypes (in blue) (data from ref. 29). Because these data are from living people, the Native American data include significant admixture from European colonization, a population with essentially no occurrence of EDAR V370A. These modern data likely vastly underrepresent the occurrence of EDAR V370A among indigenous people before European contact. Current Asian allele frequencies may be higher than pre-LGM frequencies due to back-migration from the Beringian population after the end of the LGM, a migration supported by linguistic data (46). (B) Histogram showing EDAR V370A genotype and degree of incisor shoveling, demonstrating the imperfect but strongly additive nature of EDAR V370A’s influence on incisor shoveling (adapted from ref. 43). (C) Frequencies of incisor shoveling scores observed in archaeological populations from Africa, Europe, South Asia, East Asia, North America, and South America (Table S1). Purple and blue represent a lack of incisor shoveling, and as such, an individual who is EDAR V370A−/−. Note the lack of shoveling scores of 0 and 1, and very low occurrence of 2, in the indigenous people of the Western Hemisphere, indicating a very high frequency of EDAR V370A before European contact. The post-LGM diaspora into the Americas and the subsequent centuries of European colonization (and genetic admixture) have dramatically overwritten the allelic variation of ∼20,000 y ago when EDAR V370A experienced a bout of selective pressure. To best interpret the evolutionary history of EDAR V370A, we need information on past allele frequencies. This task is difficult from a genome sequence approach because of the paucity of ancient DNA available. However, the pleiotropic effects of EDAR V370A provide a unique opportunity to more precisely reconstruct its evolutionary history without DNA sequence data. |
|
|
|
Post by Admin on Oct 27, 2019 18:26:12 GMT
Temporal and Geographic Variation of EDAR V370A As noted previously, EDAR V370A is correlated with a suite of morphological variants on teeth, the most notable of which is incisor shoveling (40⇓⇓–43). GWASs report a close relationship between genotype (EDAR V370A) and this dental phenotype in living Chinese, Japanese, and Koreans (40⇓⇓–43). In these populations, incisor shoveling of any degree correlates with the presence of one or two copies of the EDAR V370A allele in an imperfect but clearly additive manner (41⇓–43) (Fig. 3B). We scored the degree of expression in maxillary incisor shoveling for 5,333 people from >54 archaeological populations from across Europe, Asia, and North and South America (Supporting Information and Tables S1 and S2). Incisor shoveling is absent or expressed only to a slight degree outside of North and East Asian populations (where EDAR V370A frequencies are close to zero) but close to ubiquity in Native American populations, a pattern long interpreted as the result of an unknown selective pressure (47, 48) (Fig. 3C). Considering the GWAS results that indicate an additive effect of EDAR V370A’s contribution to incisor shoveling, we interpreted a score of 1 or greater as evidence of an individual likely carrying at least one copy of EDAR V370A. There are no reported instances of a score of 0 among the 3,183 individuals assessed from North and South America, and as such, no evidence of any Native Americans before European contact without at least one copy of the EDAR V370A allele (Fig. 3C and Supporting Information). A series of ANOVAs comparing the frequency of each shoveling grade (range: 0–7) across geographic regions demonstrates that a score of 1 has the greatest variation between geographic groups, further suggesting that a score of 1 or higher is reflective of the presence of at least one copy of EDAR V370A (Supporting Information and Table S3). These phenotypic data strongly suggest that incisor shoveling (and, concomitantly, the EDAR V370A allele) reached near-fixation in the population ancestral to all indigenous people of the Western Hemisphere. Given the timing of the dispersal into the Americas ∼17,000 y ago (3, 49), these dental data support the conclusion that EDAR V370A underwent positive selection in the ancestral Native American population during the LGM, similar to what we see with the FADS gene cluster (13), rather than in a population in what is now China (our evolutionary quantitative genetic analysis further supports this; Supporting Information). 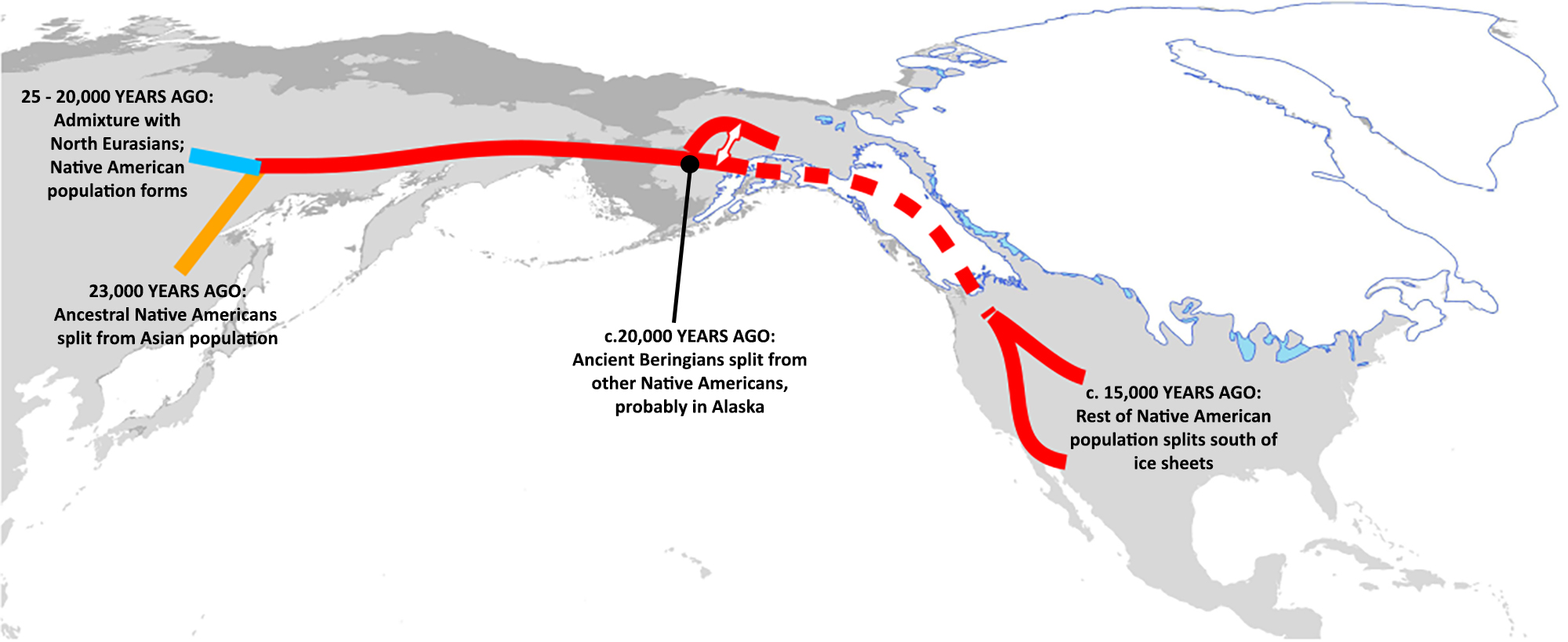 The Ancestral Population of Native Americans and the LGM Indigenous people in the Western Hemisphere derive from people who occupied latitudes above 55°N in Asia before 40,000 y ago (50). Well-dated archaeological sites near the mouth of the Yana River indicate a year-round adaptation to the Beringian Arctic Zone (latitude of 70°N) by 32,000 calibrated radiocarbon years (cal) B.P. (51). Between 28,000 and 18,000 y ago, during the LGM, as aridity increased and biological productivity reduced dramatically (52), plants and animals in many parts of the world retreated to refugia, leaving signals of genetic isolation and hybridization that are documented widely by molecular ecologists (53, 54). Human populations similarly abandoned arid regions in Africa, Eurasia, and Australia, as indicated by genetic bottlenecks and evidence of local settlement hiatus (e.g., refs. 55, 56). Various lines of evidence suggest that the Native American lineages during the LGM were genetically isolated in one or more refugia within Beringia (2, 5⇓–7), and at least one archaeological site appears to confirm a human presence in northeastern Beringia (latitude of 67°N) during that time frame (57). Analyses of Native American genetic variation indicate that the ancestral population was isolated for up to 9,000 y before dispersal in North and South America (after 15,000 cal B.P.), with an effective population size of at least a few thousand individuals (3, 49, 58). The “Beringian Standstill” model places the ancestral Native American population in the Beringian refugium during much or all of the LGM (2, 5⇓–7), where it was geographically isolated from the Siberian population by uninhabitable areas in northeastern Asia and unable to expand into the Americas because of the coalesced Laurentide and Cordilleran ice sheets (Fig. 1B). The genetic isolation is a key point, as evolution occurs more readily in the absence of gene flow (59). The FADS polymorphisms and EDAR V370A likely existed in the pre-LGM ancestral populations [although the allele is not present in Denisovans (60)]. However, there was a significant opportunity for selection to shift the frequencies of functional alleles dramatically when this population became genetically isolated in the extreme UV environment of Beringia, including Arctic Beringia (above 66°N). |
|
|
|
Post by Admin on Oct 28, 2019 18:34:18 GMT
Biological Consequences of Low UV Most vitamin D necessary for human health and reproductive success is produced through biosynthesis initiated by skin exposure to UV-B photons (61, 62). The lower rates of cutaneous synthesis of vitamin D that result from low UV environments have a wide range of health consequences. Vitamin D is among a large family of fat-soluble micronutrients that accumulate in adipose tissue proportional to circulating serum levels, and is released slowly as serum levels reduce (63). The bioactive form of vitamin D plays a secosteroid hormonal role in permitting absorption of dietary calcium through the lining of the gut (64). Through the Vitamin D receptor (Vdr), vitamin D also regulates expression of more than 220 genes in the human genome, with significant involvement in immune function and autoimmune disorders (65, 66). It is therefore not surprising that variation in immune function and disease risk correlates with latitude and UV-B exposure (67). Vitamin D also plays an important immunomodulatory role at the maternal/fetal interface (the placenta) (68) and is associated with sex-specific variation in birth weight (69). 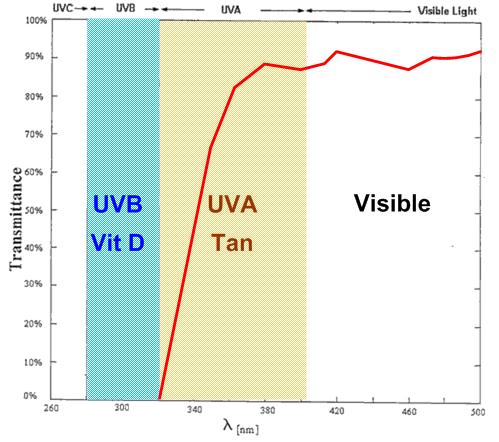 However important the immunological role of vitamin D, adipose tissue appears to be the main phenotypic target of Vdr (63, 65, 66, 70). Vitamin D deficiency significantly compromises the metabolic function of adipose tissue, with a wide range of deleterious health effects (63). Body fat has a remarkable degree of phenotypic plasticity that enables a wide range of essential physiological functions, from maintaining body heat, to responding to cold, to forming the pink adipocytes in the mammary glands during pregnancy in anticipation of lactation (71⇓–73) (Fig. 2A). Nutritional studies have long shown interrelationships between vitamin D and LC-PUFAs (74⇓–76). For example, the vitamin D synthesized through skin exposure to UV-B (vitamin D3) has positive effects on lipid profiles that cannot be replicated with dietary vitamin D2 supplementation (77). These interrelationships result from the effects of vitamin D on adipose tissue and immunological function, which are modulated by environmental exposure to UV-B. Human populations that dispersed into geographic regions above 30° latitude must have experienced selection for reduced cutaneous eumelanin pigmentation to facilitate vitamin D biosynthesis (78, 79). Selection for depigmented skin phenotypes has occurred at least twice in modern human evolution and once in Neanderthals (78, 80), with selective pressure readily detected in genomic analyses (81⇓–83).  At latitudes above 48° (e.g., 450 km north of Hokkaido, Japan; Fig. 1A), sunlight reaching the Earth’s surface contains almost no UV-B except for low levels at or near the summer solstice (84, 85). Year-round human habitation of even more extreme high latitudes leads to vitamin D deficiency that cannot be counterbalanced by depigmentation alone (84) (Fig. 1A). In these geographic regions, diets centered on vitamin D-rich foods, such as marine mammals, oily fish, reindeer, or caribou, have been adopted across a range of traditional Arctic cultures (85). Archaeological evidence shows that these dietary innovations correlate with the earliest occupation of these latitudes (55, 86, 87) and shifts in allele frequencies of the genes involved in fatty acid synthesis (16). Dietary practices can mitigate vitamin D deficiency for older children and adults, but pregnant and nursing mothers and their breast-feeding infants are still at significant risk because they need particularly high levels of vitamin D to avoid the litany of deleterious effects of vitamin D insufficiency (88, 89). |
|


 during a warm spell during the LGM (31) or that the increase in sebaceous glands would offer protection from the cold, dry air of the LGM (30). These interpretations rely primarily on the current distribution of the allele in living populations (Fig. 3A), which shows high frequencies of EDAR V370A in North and East Asian and Native American populations but virtual absence in other populations around the world (29⇓–31, 45).
during a warm spell during the LGM (31) or that the increase in sebaceous glands would offer protection from the cold, dry air of the LGM (30). These interpretations rely primarily on the current distribution of the allele in living populations (Fig. 3A), which shows high frequencies of EDAR V370A in North and East Asian and Native American populations but virtual absence in other populations around the world (29⇓–31, 45).