|
|
Post by Admin on Jun 8, 2022 19:06:26 GMT
Results Phylogenetic tree of hg N3a4 and coalescence ages of major sub-clades To reconstruct the phylogeny of hg N3a4-Z1936 and refine its inner structure we used 33 high-coverage chrY sequences, including those of 5 Hungarians (Fig. 2). All variants accumulated on the branches are listed in Supplementary Table S1, the age estimates of N3a4 sub-clades are shown on Fig. 2 and presented in Supplementary Table S2. Figure 2 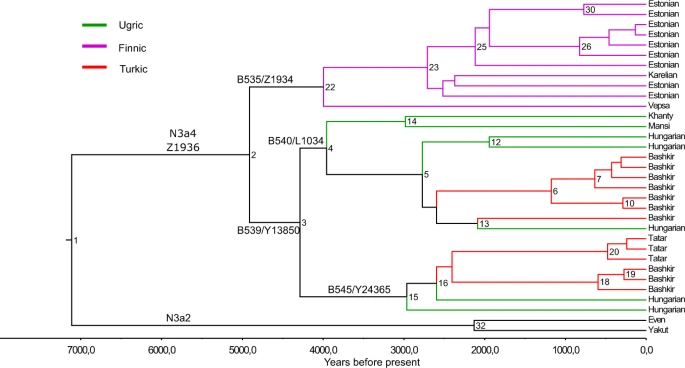 Phylogenetic tree of hg N3a4. Phylogenetic tree of 33 high coverage Y-chromosomes from haplogroup N3a4 was reconstructed with BEAST v.1.7.5 software package. We used 8 sequences published in Karmin et al.63, 6 sequences published in Ilumäe et al.27, 2 sequences published in Wong et al.64 and 17 new sequences from this study. Two N3a2 samples were used as an outgroup to estimate coalescent times. Internal node numbers on the branches (not including nodes with low posterior values), sub-clade names and population names on the tips are indicated. Branches are coloured according to language affiliations. Number of branch-defining mutations and marker names are presented in Supplementary Fig. S4. All SNPs characterizing the clades (nodes) are presented in Supplementary Table S1. Age estimates can be found in Supplementary Table S2. Phylogenetic tree of hg N3a4 has two main sub-clades defined by markers B535 and B539 that diverged around 4.9 kya (95% confidence interval [CI] = 3.7–6.3 kya). Inner sub-clades of N3a4-B539 (defined by markers B540 and B545) split 4.2 kya (95% CI = 3.0–5.6 kya). Further sub-clades of B540, one containing Y-chromosomes of a Khanty and a Mansi and other containing chrY’s of Hungarians and Bashkirs (Fig. 2, Supplementary Table S2) split around 3.9 kya (95% CI = 2.8–5.2 kya). The expansions of B540 and B545 started around the same time about 2.7–2.9 kya (B540-PH573/L1442: 95% CI = 1.8–3.7 kya; B540-Y28538: 95% CI = 2.0–4.1 kya; B545: 95% CI = 2.0–4.1 kya) (Supplementary Table S2). The phylogenetic tree reveals that all five Hungarian samples belong to N3a4-B539 sub-clade that they share with Ob-Ugric speaking Khanty and Mansi, and Turkic speaking Bashkirs and Tatars from the Volga-Ural region. Hungarian and Bashkir chrY lineages belong to both sub-clades of N3a4-B539 (Fig. 2). |
|
|
|
Post by Admin on Jun 9, 2022 18:04:28 GMT
Geographic distribution of genotyped hg N sub-lineages Although, the frequency of hg N among modern Hungarians is only marginal, this small, but very intriguing from historical point of view, portion of paternal lineages belongs to N3a4 clade. To test the presence and proportions of hg N3a4 lineages in a more comprehensive sample set and with a higher phylogenetic resolution level compared to earlier studies14,15,16,31, we analysed the genotyping data of about 5000 Eurasian individuals, including West Siberian Mansi and Khanty who are linguistically closest to Hungarians (Supplementary Fig. S1). Genotyping confirms that the N3a4-B539 clade found among Hungarians, Bashkirs and Tatars is also shared with Ugric speaking Mansis and Khantys (Supplementary Table S3), matching the findings of Feher et al.31 and showing that B539 clade is common for all Ugric speaking populations. To visualize the geographic spread of hg N3a4 and its sub-clades we used genotype frequencies (Supplementary Tables S3 and S4) to construct distribution maps (Fig. 3). Figure 3 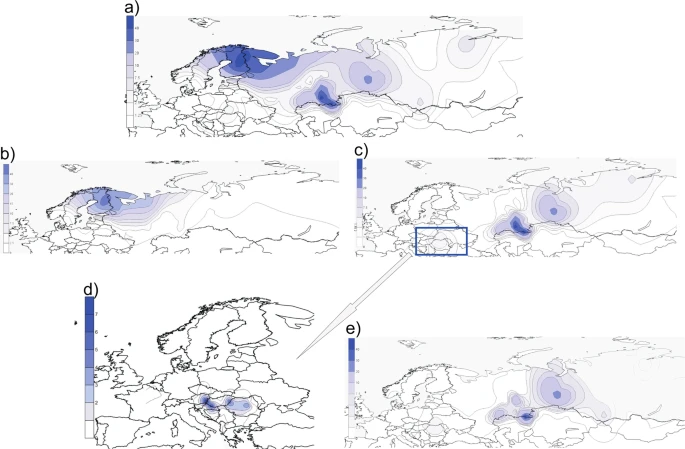 Frequency(%) distribution maps of individual sub-clades of hg N3a4. (a) Map of the entire hg N3a4. (b,c) Maps of N3a4 sub-clades defined by B535 and B539, respectively. (d) The local snapshot of B539 is a subsample of points presented on panel (c) showing the N3a4-B539 distribution among Hungarian speakers. Note that the scales of the maps are different. (e) Map of N3a4-B540/L1034. All the maps were created with Surfer v.8. Data points used for generating the maps are given in Supplementary Tables S3 and S4 for local B539 map. Background maps from Surfer® (v.8, Golden Software, Inc, Golden, CO, USA). N3a4 has two frequency peaks (Fig. 3a) – one in Northeast Europe where mostly N3a4-B535 is found (Fig. 3b), and the other in southern slopes of the Ural Mountains where N3a4-B539 is prevalent (Fig. 3c). There is a clear difference in geographic distribution patterns of these two hg N3a4 sub-clades. Hg N3a4-B535 (Fig. 3b) is common mostly among Finnic (Finns, Karelians, Vepsas, Estonians) and Saami speaking populations in North eastern Europe (Supplementary Table S3). The highest frequency is detected in Finns (~44%) but it also reaches up to 32% in Vepsas and around 20% in Karelians, Saamis and North Russians. The latter are known to have changed their language or to be an admixed population with reported similar genetic composition to their Finnic speaking neighbors36. The frequency of N3a4-B535 rapidly decreases towards south to around 5% in Estonians, being almost absent in Latvians (1%) and not found among Lithuanians. Towards east its frequency is from 1–9% among Eastern European Russians and populations of the Volga-Ural region such as Komis, Mordvins and Chuvashes, but it is absent among Tatars and Bashkirs with an exception of Burzyansky District Bashkirs where the frequency is up to 5% (Supplementary Table S3). Hg N3a4-B539 (Fig. 3c), on the other hand, is prevalent among Turkic speaking Bashkirs and also found in Tatars but is entirely missing from other populations of the Volga-Ural region such as Uralic speaking Udmurts, Maris, Komis and Mordvins, and in Northeast Europe, where instead N3a4-B535 lineages are frequent. Besides Bashkirs and Tatars in Volga-Ural region, N3a4-B539 is substantially represented in West Siberia among Ugric speaking Mansis and Khantys. Among Hungarians, however, N3a4-B539 has a subtle frequency of 1–4% (Fig. 3d), which is surprising considering their distant location from the core area of N3a4-B539. The sub-clades of N3a4-B539 (B540 and B545) (Fig. 2) have partly overlapping distribution areas with highest frequency peaks of hg N3a4-B540/L1034 (Fig. 3e) in the Ural Mountain region. B540 is more widespread and the most frequent among different sub-populations of Bashkirs in Southern Urals (up to 60%) and in West Siberia among Ugric speaking Mansis and Khantys (up to 27%). It is also present in Uralic speaking Nenets (7%) and Turkic speaking Dolgans (5%) but missing from Uralic speaking Nganasans and Selkups. Although N3a4-B540 is prevalent in Bashkirs it has a considerably lower frequency among neighbouring Tatars (3–5%). B540/L1034 sister-clade B545, however, has a much more localised distribution area – it is present with high frequencies (52%) in Volga-Ural region only among Bashkirs from Sterlibashevsky district of Bashkortostan (Supplementary Table S3). Despite the low sample size, it is notable that both 10th century ancient Hungarian samples from Upper-Tisza region that were derived at Z1936 also belong to the B545 clade32. Additionally, contemporary Mansis, who have a high amount of N3a4-B540, lack N3a4-B545. Most intriguingly, similarly to Bashkirs and Tatars both N3a4-B539 sub-clades can be found among Hungarians (Fig. 2, Supplementary Table S3). The frequency of B540 lineage is highest among Sekler Hungarians (around 4%), who today live in Transylvania, Romania. The Sekler are half-million Hungarian speaking people whose historical background is not completely understood. They are distinguished from other Hungarian groups even in the earliest Hungarian chronicles37 and form a geographically and genetically isolated sub-population among Hungarians. Csanyi et al.15 has shown earlier that Sekler Hungarians have hg N3 (1%), but the resolution level used in this study does not allow us to specify further sub-clades. In other Hungarian sub-groups, the frequency of the B540 clade is around 1%. The B545-clade is absent in Sekler Hungarians and is less than 1% in other Hungarian sub-groups (Supplementary Table S3). |
|
|
|
Post by Admin on Jun 9, 2022 21:59:44 GMT
To further reveal the inner structure within N3a4 sub-clades we constructed a median-joining network for all Z1936 carrying Y-chromosomes based on 16 STR loci (Supplementary Fig. S2, Supplementary Table S4). The N3a4 network shows a clear clustering of sub-haplogroups. The B535 sub-clade consists of a loose cluster prevalent among Vepsas and Central Russians and a star-like cluster that contains all Finnish, Saami, Northern Russians, Mordovin, as well as the overwhelming majority of Estonian and Karelian samples plus one Ukrainian and Vepsa sample. Such STR pattern indicates a possible SNP-based sub-structure of the clade. The B540/L1034 sub-cluster has a distinguished star-like pattern with a major founder haplotype containing 14 STR haplotypes from different studied populations; Bashkirs, Mansis, Hungarians, Khanty and Tatars. The three Ukrainian, the single Greek and a West Hungarian sample represent a smaller branch. The B545 network did not form a clear sub-cluster in the network, this might be due to the relatively low number of available haplotypes.
Two ancient DNA samples of Hungarian Conquerors derived at Z193632, were included in the network. Their B545 status could not be confirmed, but their STR pattern showed a closer relationship to contemporary Bashkir and Tatar samples than present-day Hungarians.
We tested whether a simple scenario with no gene flow nor admixture can explain the observed frequencies of haplogroup N3a4-B539 in Hungarian Sekler (4%), their neighbouring Indo-European populations (0%) and Southern Ural/West Siberian populations (13%). For that we implemented 3 models: A, B and C (see Material and Methods, Supplementary Fig. S5). Our simplified model shows that Model A depicting the situation with drift alone is almost never accepted (mean 0.04% and standard deviation 0.19%). Although Model B, where drift affects Hungarian Seklers and Indo-Europeans after receiving same external genetic contribution, is more accepted (mean 0.58% and standard deviation 0.73%), it is still significantly rejected (more than 95% of the time). Thus, the other alternative model C, where Southern Ural/West Siberian populations have different contributions in Hungarian and their neighbouring Indo-European populations, is accepted. In all the models we assume that the contribution of N3a4-B539 is coming from Southern Ural/West Siberian populations and the expected frequency of the carriers of the B539 among them is 12.4% (6.8–18% Confidence Interval [CI]; Supplementary Fig. S3). According to Model C the expected contribution of Southern Ural/West Siberian populations to Hungarian populations is 43% (0–89% CI), but the wide CI suggest that we do not have much power in this frequency estimate. Whereas we have much higher confidence in estimating the negligible 3.3% (0–10% CI) contribution of B539 in Indo-European populations.
|
|
|
|
Post by Admin on Jun 11, 2022 17:37:49 GMT
Discussion The comparison of genetic and cultural history of human populations has excited scientists for decades10,35,38,39,40,41,42,43,44. It is widely accepted among both archaeologists and linguists that the earlier (pre)historical phase of Hungarians points to West Siberia, East of the Ural Mountains6,7,8,9,45,46,47,48 (Fig. 1a), but there is a debate about the potential archaeological cultures involved3,46 (Fig. 1b). The first widely accepted station on the early Hungarian migration route is the Kushnarenkovo (6th‒8th centuries CE)49,50,51,52 and the succeeding Karayakupovo cultures (9th‒10th centuries CE)52,53,54, with sites mainly found in present-day Bashkortostan, Tatarstan and in the Chelyabinsk area of the Trans-Ural region8. Based on the archaeological evidence, a portion of the Hungarians moved to the west in the middle of the 9th century and appeared near the lower reaches of the Dnieper River in present-day Ukraine, where Hungarians are mentioned in historical sources8. However, archaeological sites of Chiyalikskaya culture (11th–13th) support the survival of Hungarians in the Ural region55,56. Furthermore, other sources confirmed the survival of Hungarians in the Ural region until the 13th century45,46. Also, the eastern Hungarian homeland called Magna Hungaria (Great or Ancient Hungary) is referred in the early Hungarian chronicles48,57. Studies based on chrY have shown that the frequency spectrum of different chrY haplogroups and sub-clades varies in large range in different sub-populations58, a phenomenon observable also in our study of N3a4 clade (ranging in frequency from 1–59%) (Supplementary Table S3). Compared to earlier study by Feher et al.31, where shared lineages of hg N3a4-Z1936 between geographically distant but linguistically close Hungarians and Mansi were first reported, present study covers more populations and a wider geographical area (Supplementary Table S3). While chrY hgs usually show smooth distribution patterns, the particular spatial distribution of hg N3a4-B539 and its sub-clades is important in distinguishing the paternal roots of Hungarians. The eastern roots of hg N have been revealed earlier by comprehensive phylogenetic and phylogeographic study of contemporary hg N lineages by Ilumäe et al.27. Different hg N lineages among aDNA samples from East Asia and Baikal region show that hg N was frequent and diverse in Neolithic China59 and Baikal region60 already more than 6000 years ago. N3a4-B539 sub-clades have a well-centred frequency cline in Ural region and West Siberia, but the presence of these clades in only among geographically distant Hungarians and not among any of their neighbours (Fig. 3, Supplementary Table S5) is noteworthy. Simulations enable us to reject a simple random drift model and a single migration model between all Europeans and Southern Ural/West Siberian populations. We also show that there is clearly a direct higher contribution from Southern Urals/West Siberia to Hungarian populations, but the amount cannot be pinpointed using our current data (Supplementary Fig. S3). This might reflect the migration of ancestors of Hungarians from the Ural region to the Carpathian Basin, that is also documented in historical records. The occurrence of hg N3a4-Z1936 among the remains of the individuals from the archaeologically richest 10th century cemeteries of the Hungarian Conquerors in the Carpathian Basin lend support to the Ural region origin of at least part of the Hungarian Conquerors. It is important to note that there are two different N3a-L708 subgroups that are frequent around the Baltic Sea with a clear south-north frequency gradient tendency27. N3a3-VL29 is frequent among Estonians and Latvians and can be detected also among Ukrainians, being the most westward distributed sub-clade among N3a27 and therefore finding it in low frequency (0–4%) at the outer borders of distribution zone in different Hungarian sub-populations is expected (Supplementary Table S3). The case of N3a4-B539 and its sub-clades is different: presence of these clades in the Volga-Ural region, in West Siberia and in geographically distant Hungarians is not so easy to explain by gradual frequency cline and without assuming a migration of people (who might have been among the ancestors of the present-day Hungarians). The split between of B540 and B545 subgroups within Baskhirian, Tatar and Hungarian populations started around 2700–2900 yBP (Fig. 2, Supplementary Table S2) that is in accordance with the recent linguistic data about the divergence of Ob-Ugric and Hungarian languages61. It has been proposed that the ‘Ugric Age’ lasted at least until the late Bronze Age in West Siberia and the split between Ob-Ugric and Hungarian from the common proto-Ugric branch of the Uralic language tree occurred during the first centuries of the first millennium BC9, but the recent linguistic reconstructions of the Uralic language tree give much broader borders for the divergence of Ugric clade (4900‒1700 yBP)61. The time-frame is the same for the cooling climate in West Siberia with its peak at the 9th and 8th CE which could have resulted in the movements of several West Siberian populations61,62. Although the frequency of hg N3a4-B539 is subtle among present-day Hungarians, it is possible that ancient Magyars who lived in the Ural Mountain region had a significantly higher proportion of chrY hg N, since the Z1936 lineage was found from 5 individuals out of 19 (26,3%) in the archaeologically richest Hungarian late 9th-early 10th century cemeteries32. This frequency is quite similar to Z1936 > B539 frequencies found among various Khanty, Mansi and Bashkir groups (Supplementary Table S3). The homeland of ancient Hungarians around the Ural Mountain region, and the Hungarian affinities of Kushnarenkovo and Karayakupovo cultures is widely accepted among researchers47,49,50,51,52,53,54. Further studies of chrY and autosomal diversity in ancient samples of the representatives of those cultures could also provide new insight into the demographic history of the Hungarians. Supplementary information Supplementary informationSupplementary Table S1 static-content.springer.com/esm/art%3A10.1038%2Fs41598-019-44272-6/MediaObjects/41598_2019_44272_MOESM2_ESM.xlsxSupplementary Table S2 static-content.springer.com/esm/art%3A10.1038%2Fs41598-019-44272-6/MediaObjects/41598_2019_44272_MOESM3_ESM.xlsxSupplementary Table S3 static-content.springer.com/esm/art%3A10.1038%2Fs41598-019-44272-6/MediaObjects/41598_2019_44272_MOESM4_ESM.xlsxSupplementary Table S4 static-content.springer.com/esm/art%3A10.1038%2Fs41598-019-44272-6/MediaObjects/41598_2019_44272_MOESM5_ESM.xlsxSupplementary Table S5 static-content.springer.com/esm/art%3A10.1038%2Fs41598-019-44272-6/MediaObjects/41598_2019_44272_MOESM6_ESM.xlsxSupplementary Table S6 static-content.springer.com/esm/art%3A10.1038%2Fs41598-019-44272-6/MediaObjects/41598_2019_44272_MOESM7_ESM.xlsxSupplementary Table S7 static-content.springer.com/esm/art%3A10.1038%2Fs41598-019-44272-6/MediaObjects/41598_2019_44272_MOESM8_ESM.xlsx |
|

