|
|
Post by Admin on Apr 7, 2021 22:53:52 GMT
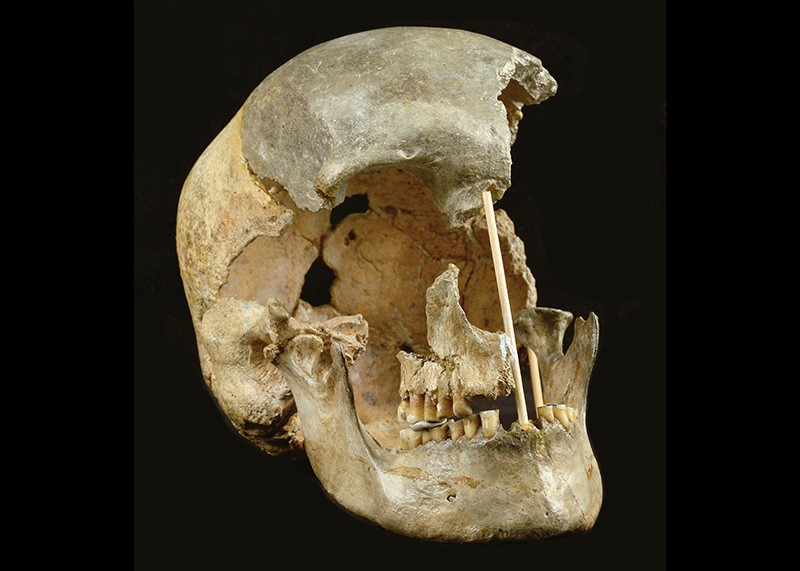 Scientists have sequenced the oldest Homo sapiens DNA on record, showing that many of Europe’s first humans had Neanderthals in their family trees. Yet these individuals are not related to later Europeans, according to two genome studies of remains dating back more than 45,000 years from caves in Bulgaria and the Czech Republic1,2. The research adds to growing evidence that modern humans mixed regularly with Neanderthals and other extinct relatives, says Viviane Slon, a palaeogeneticist at the University of Tel Aviv in Israel. “It’s different times, different places, and it happens again and again.” The genetic history of the earliest humans in Europe and Asia has been blurred. Although researchers have sequenced DNA from Neanderthals and other extinct human relatives dating as far back as 430,000 years, there is a scarcity of genetic information from the period between around 47,000 and 40,000 years ago, known as the Initial Upper Palaeolithic, and no Homo sapiens DNA at all from before this period. Genomes belonging to humans from Siberia and Romania showed no connection to later waves of Europeans, but a 40,000-year-old individual from China is a partial ancestor of modern East Asian people. Like all present-day people whose ancestry isn’t solely African, these early Eurasians carried Neanderthal DNA. Researchers thought that probably originated from mixing between the groups in the Middle East 50,000–60,000 years ago. But a 2015 study3 of the genome of the 40,000-year-old Romanian individual, from a site called Peștera cu Oase, held a surprise: a Neanderthal ancestor in the past four to six generations, suggesting that humans interbred with Neanderthals in Europe, too. It was not clear from the Oase man’s genome whether interbreeding was common in Europe. He lived around the time when Neanderthal populations, already sparse, were beginning to vanish from the region. Genetic mixture The latest genome studies, both published on 7 April, clarify the relationships between Europe’s first modern humans, later Homo sapiens and Neanderthals, but also throw up some new questions. One study, in Nature1, is based on a tooth and fragmentary remains from Bacho Kiro Cave in Bulgaria; the other, in Nature Ecology and Evolution2, looks at a nearly complete skull from a cave known as Zlatý kůň in the Czech Republic. The three oldest Bacho Kiro individuals, dated to between 45,900 and 42,600 years old, all had recent Neanderthal forebears, reports a team led by molecular biologist Mateja Hajdinjak and evolutionary geneticist Svante Pääbo, both at the Max Planck Institute for Evolutionary Anthropology (MPI–EVA) in Leipzig, Germany. The genomes of modern non-Africans usually harbour about 2% Neanderthal ancestry, but the Bacho Kiro individuals had slightly more: 3.4-3.8%, and the chromosome segments — which shorten in successive generations — were considerably longer. By measuring these segments, the researchers estimated that the Bacho Kiro individuals had Neanderthal ancestors as recently as the past six or seven generations — and probably in Europe, not the Middle East. “We saw these huge chunks. It was completely amazing,” says Hajdinjak, who is now at the Francis Crick Institute in London and was part of the team that identified the same patterns in the Oase man’s genome. “What are the chances of finding them again?” The Zlatý kůň woman’s Neanderthal ancestry goes back considerably longer: 70–80 generations, or perhaps 2,000–3,000 years, says Johannes Krause, a palaeogeneticist at the MPI–EVA who co-led the study. His team could not date the skull accurately because of contamination. But on the basis of its Neanderthal ancestry, Krause suspects it is well over 45,000 years old, and in the same ballpark as the oldest remains from Bacho Kiro. “We do have, now, some of the oldest human genomes out there,” adds Hajdinjak. Tracing the lineage The oldest individuals from Bacho Kiro and the Zlatý kůň female are not related to later Europeans, ancient or modern, meaning that their lineages must have disappeared from the region. But, to their surprise, Hajdinjak and her colleagues found that the Bacho Kiro people shared a connection with contemporary East Asians and Native Americans. Hajdinjak suggests that the Bacho Kiro remains represent a population that once lived across Eurasia, but vanished from Europe and lived on in Asia. The fact that several humans from Bacho Kiro had very recent Neanderthal relatives suggests that the groups mixed routinely in Europe, says Marie Soressi, an archaeologist at the University of Leiden in the Netherlands who plans to examine European archaeology through this lens. Stone tools and other artefacts common to the Initial Upper Palaeolithic — and distinct from typical Neanderthal and later human toolkits — could be a product of cultural exchanges or even mixed populations, she says. “We really want to better understand what happened, what was the historical process and how peaceful were those encounters.” doi: doi.org/10.1038/d41586-021-00916-0References 1.Hajdinjak, M. et al. Nature doi.org/10.1038/s41586-021-03335-3 (2021). 2.Prüfer, K. et al. Nature Ecol. Evol. doi.org/10.1038/s41559-021-01443-x (2021). 3.Fu, Q. et al. Nature 524, 216–219 (2015). |
|
|
|
Post by Admin on May 3, 2022 17:37:52 GMT
Initial Upper Palaeolithic humans in Europe had recent Neanderthal ancestry Mateja Hajdinjak, Fabrizio Mafessoni, …Svante Pääbo Nature volume 592, 253–257 (2021) Abstract Modern humans appeared in Europe by at least 45,000 years ago1,2,3,4,5, but the extent of their interactions with Neanderthals, who disappeared by about 40,000 years ago6, and their relationship to the broader expansion of modern humans outside Africa are poorly understood. Here we present genome-wide data from three individuals dated to between 45,930 and 42,580 years ago from Bacho Kiro Cave, Bulgaria1,2. They are the earliest Late Pleistocene modern humans known to have been recovered in Europe so far, and were found in association with an Initial Upper Palaeolithic artefact assemblage. Unlike two previously studied individuals of similar ages from Romania7 and Siberia8 who did not contribute detectably to later populations, these individuals are more closely related to present-day and ancient populations in East Asia and the Americas than to later west Eurasian populations. This indicates that they belonged to a modern human migration into Europe that was not previously known from the genetic record, and provides evidence that there was at least some continuity between the earliest modern humans in Europe and later people in Eurasia. Moreover, we find that all three individuals had Neanderthal ancestors a few generations back in their family history, confirming that the first European modern humans mixed with Neanderthals and suggesting that such mixing could have been common. Main The transition between the Middle and Upper Palaeolithic periods in Europe, which started about 47,000 years before present (47 kyr BP)1,2, overlapped with the spread of modern humans and the disappearance of Neanderthals, which occurred by about 40 kyr BP6. Analyses of the genomes of Neanderthals and modern humans have shown that gene flow occurred between the two hominin groups approximately 60–50 kyr BP8,9,10,11, probably in southwestern Asia. However, owing to the scarcity of modern human remains from Eurasia that are older than 40 kyr1,2,3,4,5,12, genome-wide data are available for only three individuals of this age7,8,13 (Fig. 1). Little is therefore known about the genetics of the earliest modern humans in Eurasia, the extent to which they interacted with archaic humans and their contribution to later populations. For example, whereas the roughly 42,000 to 37,000-year-old ‘Oase1’ individual from Romania7,14 and the roughly 45,000-year-old ‘Ust’Ishim’ individual from Siberia8 do not show specific genetic relationships to subsequent Eurasian populations, the approximately 40,000-year-old ‘Tianyuan’ individual from China contributed to the genetic ancestry of ancient and present-day East Asian populations13. Another open question is the extent to which modern humans mixed with Neanderthals when they spread across Europe and Asia. Direct evidence of local interbreeding exists only for the Oase1 individual, who had a recent Neanderthal ancestor7 in his family history. 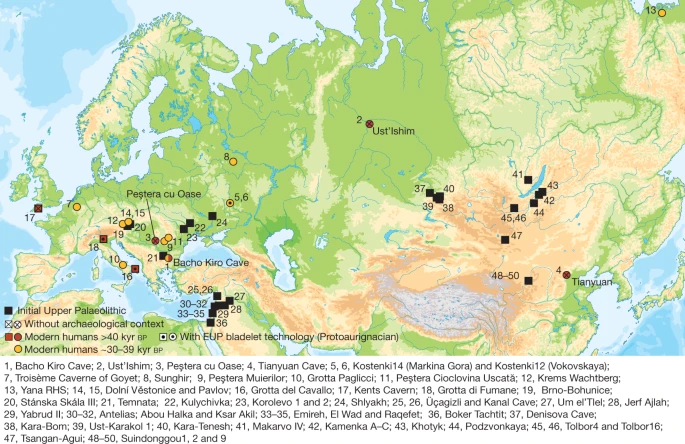 Fig. 1: Archaeological sites that have yielded genetic data and/or IUP assemblages. Sites with modern human genome-wide data older than 40 kyr BP (red circles) or older than 30 kyr BP (yellow circles), sites in Europe with modern human remains older than 40 kyr BP (red squares) and sites with IUP assemblages (black squares). |
|
|
|
Post by Admin on May 3, 2022 18:47:14 GMT
Here, we analyse genome-wide data from human specimens found in direct association with an Initial Upper Palaeolithic (IUP) assemblage of artefacts in Bacho Kiro Cave, Bulgaria1 (Fig. 1), as well as from two more recent specimens from the same site (Supplementary Information 1). The IUP groups together assemblages that fall chronologically between the last Middle Palaeolithic assemblages and the first bladelet industries of the Upper Palaeolithic. The IUP spans a broad geographical area15, from southwest Asia, central and eastern Europe to Mongolia16 (Fig. 1, Supplementary Information 1). Although there are reasons to group these assemblages on the basis of their lithic technology, the IUP also shows great regional variability. Therefore, it is debated whether the IUP represents a dispersal of modern humans across middle-latitude Eurasia, the diffusion of certain technological ideas, instances of independent invention, or a combination of some or all of these15. The IUP is contemporaneous with late Neanderthal sites in central and western Europe6 and precedes later Upper Palaeolithic techno-complexes in Europe, such as the Protoaurignacian and the Aurignacian, by several thousand years5.
Five human specimens were recovered from Bacho Kiro Cave in recent excavations. They consist of a lower molar (F6-620) found in the upper part of Layer J in the Main Sector, and four bone fragments (AA7-738, BB7-240, CC7-2289 and CC7-335) from Layer I in Niche 1. They have been directly radiocarbon-dated to between 45,930 and 42,580 calibrated years before present (cal. BP)1,2, and their mitochondrial genomes are of the modern human type, suggesting that they are the oldest Upper Palaeolithic modern humans that have been recovered in Europe1. One bone fragment was found in Layer B in the Main Sector (F6-597) and another one was among the finds from excavations in the 1970s, when it was retrieved in a position corresponding to the interface of Layers B and C (BK1653). The two latter bone fragments were directly dated to 36,320–35,600 cal. BP and 35,290–34,610 cal. BP1,2, respectively. Although the lithic assemblages from the later layers are sparse, they are likely to be Aurignacian1,2. We also produced additional data from a mandible7,14 that was found outside any archaeological context in Peștera cu Oase, Romania (referred to as ‘Oase1’)14. The mandible was directly dated to about 42–37 kyr BP14, although this may be an underestimate as the dating was performed before recent technical improvements.
We extracted DNA from between 29.3 mg and 64.7 mg of tooth or bone powder from the specimens as described1. We also treated 15 mg of bone powder from the Oase1 mandible with 0.5% hypochlorite solution to reduce bacterial and human contamination before DNA extraction17. Among DNA fragments sequenced from the DNA libraries constructed from the Bacho Kiro Cave and Oase1 extracts, between 0.003% and 1.8% could be mapped to the human genome (Supplementary Information 2). Owing to the low fraction of hominin DNA, we used in-solution hybridization capture18 to enrich the libraries for about 3.8 million single-nucleotide polymorphisms (SNPs) that are informative about modern human variation and archaic admixture7,19 (excluding F6-597, which contained very little if any endogenous DNA; Supplementary Information 2).
For the six specimens, between 57,293 and 3,272,827 of the targeted SNPs were covered by at least one DNA fragment (Extended Data Table 1). Of these, between 11,655 and 2,290,237 SNPs were covered by at least one fragment showing C-to-T substitutions in the first three and/or the last three positions from the ends, suggesting the presence of deaminated cytosine bases, which are typical of ancient DNA20 (Extended Data Table 1, Extended Data Fig. 1). On the basis of the numbers of putatively deaminated fragments aligning to the X chromosome and the autosomes21 (Supplementary Information 4), we conclude that specimens F6-620, AA7-738, BB7-240 and CC7-335 belonged to males, whereas BK1653 and CC7-2289 belonged to females, although the low amount of data makes this conclusion tentative for CC7-2289 (Extended Data Fig. 2a).
Using an approach that makes use of DNA deamination patterns22, we estimated that the overall nuclear DNA contamination was between 2.2% ± 0.5% (F6-620) and 42.4% ± 0.6% (CC7-2289). In the male specimens, we estimated contamination from polymorphisms on the X chromosome23 to between 1.6% ± 0.1% and 3.4% ± 0.5% (Supplementary Information 2). Owing to the presence of present-day human contamination, we restricted all downstream analyses to putatively deaminated fragments for all specimens except F6-620 (for which contamination was so low that we used all fragments). This left between 11,655 and 3,272,827 SNPs per specimen to be used for the subsequent analyses (Supplementary Information 2).
The molar F6-620 and the bone fragment AA7-738 have identical mitochondrial genome sequences1 and both come from males. The pairwise mismatch rate between the two specimens at the SNPs24 is 0.13, similar to the mismatch rate between libraries from the same specimen (Extended Data Fig. 2b). By contrast, this number is 0.23 (interquartile range: 0.22–0.25) for the other Bacho Kiro Cave specimens, similar to unrelated ancient individuals from other studies (Extended Data Fig. 2b). Thus, we conclude that specimens F6-620 and AA7-738 belonged to the same individual or to identical twins, which is much less likely.
We enriched the libraries from the male individuals using probes that targeted about 6.9 Mb of the Y chromosome25 (Supplementary Information 4) and arrived at 15.2-fold coverage for F6-620, 2.5-fold for BB7-240 and 1.5-fold for CC7-335. F6-620 carries a basal lineage of the Y chromosome haplogroup F (F-M89), whereas BB7-240 and CC7-335 carry haplogroup C1 (C-F3393). Although haplogroup C is common among males from East Asia and Oceania, both haplogroups F and C1 are rare in present-day humans and are found only at low frequencies in mainland Southeast Asia and Japan26,27.
We estimated the extent of genetic similarity among the Bacho Kiro Cave individuals and other early modern humans using outgroup f3-statistics28. The three roughly 45,000-year-old IUP individuals are more similar to one another than to any other ancient individual (Extended Data Fig. 3a). By contrast, BK1653, which is about 35,000 years old, is more similar to later Upper Palaeolithic individuals from Europe who are around 38,000 years old or younger29,30 (3.0 ≤ |Z| ≤ 17.4; Extended Data Fig. 4, Supplementary Information 5); for example, to the roughly 35,000-year-old ‘GoyetQ116-1’ individual from Belgium and members of the ‘Věstonice’ genetic cluster, who are associated with later Gravettian assemblages29 (Extended Data Figs. 3a, 4b, c).
|
|
|
|
Post by Admin on May 3, 2022 20:25:39 GMT
When comparing the Bacho Kiro Cave individuals to present-day populations31, we found that the IUP individuals share more alleles (that is, more genetic variants) with present-day populations from East Asia, Central Asia and the Americas than with populations from western Eurasia (Fig. 2a, Supplementary Information 5), whereas the later BK1653 individual shares more alleles with present-day western Eurasian populations (Extended Data Figs. 3b, 4a). Fig. 2: Population affinities of the IUP Bacho Kiro Cave individuals. 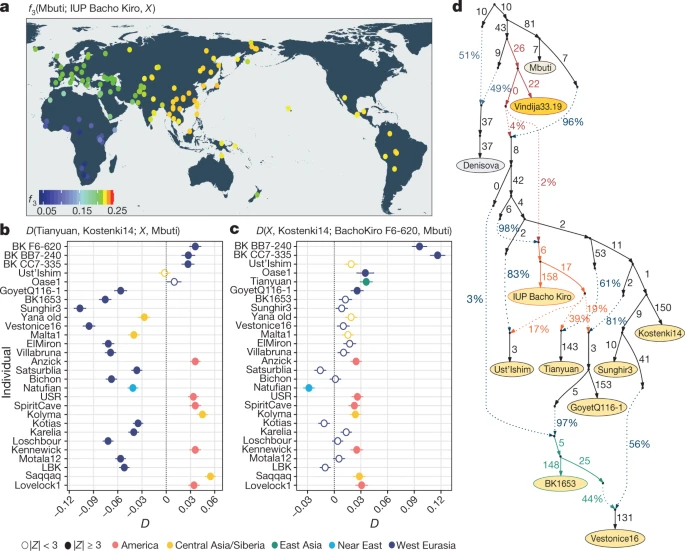 a, Allele sharing (f3) between the IUP Bacho Kiro Cave individuals and present-day populations (X) from the Simons Genome Diversity Project (SGDP)31 after their separation from an outgroup (Mbuti) (calculated as f3(Mbuti; IUP Bacho Kiro, X). Warmer colours on the map48 correspond to higher f3 values (higher shared genetic drift). b, IUP Bacho Kiro Cave individuals share significantly more alleles (proportions of allele sharing or D values plotted on x axis) with the roughly 40,000-year-old Tianyuan individual13 than with the approximately 38,000-year-old Kostenki14 individual29,30. Calculated as D(Tianyuan, Kostenki14; X, Mbuti). c, F6-620 shares significantly more alleles with the Oase17 and GoyetQ116-129 individuals, ancient Siberians and Native American individuals than with the Kostenki14 individual. Calculated as D(X, Kostenki14; F6-620, Mbuti). b, c, Filled circles indicate a significant value (|Z| ≥ 3); open circles, |Z| < 3. Whiskers correspond to 1 s.e. calculated across all autosomes (1,813,821 SNPs) using a weighted block jackknife28 and a block size of 5 Mb. BK, Bacho Kiro. d, Admixture graph relating Bacho Kiro Cave individuals and ancient humans older than 30 kyr BP. This model uses 281,732 overlapping SNPs in all individuals and fits the data with a single outlier (Z = 3.22). Ancient non-Africans (yellow circles), Vindija 33.19 Neanderthal (orange), Denisovan (grey) and present-day African individuals (light yellow circle) are shown. Admixture edges (dotted lines) show the genetic component related to Neanderthals (red), to the IUP Bacho Kiro Cave individuals (orange) and to BK1653 (green). Numbers on solid branches correspond to the estimated drift in f2 units of squared frequency difference; labels on dotted edges give admixture proportions. We next investigated whether these observations could be due to the fact that present-day populations in western Eurasia derive part of their ancestry from ‘Basal Eurasians’32,33, an inferred population that diverged early from other non-African populations and may have ‘diluted’ allele sharing between western Eurasian populations and IUP individuals. To do this, we compared the Ust’Ishim, Oase1 and IUP Bacho Kiro Cave individuals to western Eurasian individuals such as the approximately 38,000-year-old ‘Kostenki14’ individual from Russia29,30, which pre-dates the introduction of ‘Basal Eurasian’ ancestry to Europe around 8,000 cal. BP32. We found that the Ust’Ishim and Oase1 individuals showed no more affinity to western than to eastern Eurasian populations, suggesting that they did not contribute ancestry to later Eurasian populations, as previously shown7,8 (Supplementary Information 5, Extended Data Fig. 5). By contrast, the IUP Bacho Kiro Cave individuals shared more alleles with the roughly 40,000-year-old Tianyuan individual13 from China (Fig. 2b) and other ancient Siberians34,35 and Native Americans36,37,38,39 (Fig. 2c) than with the Kostenki14 individual (3.6 ≤ |Z| ≤ 5.3). Among other western Eurasian Upper Palaeolithic humans, the IUP Bacho Kiro Cave individuals shared more alleles with the Oase1 (3.6 ≤ |Z| ≤ 4.3) and roughly 35,000-year-old GoyetQ116-129 individuals than with the Kostenki14 individual (3.2 ≤ |Z| ≤ 4.3; Fig. 2c, Supplementary Information 5). Notably, the GoyetQ116-1 individual has previously been shown to share more alleles with early East Asians than other individuals of a similar age in Europe13. When we explored models of population history that are compatible with the observations above using admixture graphs28, we found that the IUP Bacho Kiro Cave individuals were related to populations that contributed ancestry to the Tianyuan individual in China as well as, to a lesser extent, to the GoyetQ116-1 and Ust’Ishim individuals (all |Z| < 3; Fig. 2d, Supplementary Information 6). This resolves the previously unclear relationship between the GoyetQ116-1 and Tianyuan individuals13 without the need for gene flow between these two geographically distant individuals. The models also suggest that the later BK1653 individual belonged to a population that was related, but not identical, to that of the GoyetQ116-1 individual (Fig. 2d, Extended Data Fig. 4, Supplementary Information 6) and that the Věstonice cluster, whose members were found in association with Gravettian assemblages29, derived part of their ancestry from such a population and the rest from populations related to the roughly 34,000-year-old ‘Sunghir’ individuals40 from Russia (Fig. 2d, Supplementary Information 6). As the IUP Bacho Kiro Cave individuals lived at the same time as some of the last Neanderthals in Europe6, we estimated the proportion of Neanderthal DNA in their genomes by taking advantage of two high-quality Neanderthal genomes9,10,41. We found that the IUP individuals F6-620, BB7-240 and CC7-335 carried 3.8% (95% confidence interval (CI): 3.3–4.4%), 3.0% (95% CI: 2.4–3.6%) and 3.4% (95% CI: 2.8–4.0%) Neanderthal DNA, respectively. This is more than the average of 1.9% (95% CI: 1.5–2.4%) found in other ancient or present-day humans, except for the Oase1 individual, who had a close Neanderthal relative (6.4% (95% CI: 5.7–7.1%); Extended Data Fig. 6, Supplementary Information 7). By contrast, the more recent BK1653 individual carried only 1.9% (95% CI: 1.4–2.4%) Neanderthal DNA, similar to other ancient and present-day humans10,41 (Extended Data Fig. 6). As has been the case for all humans studied so far, the Neanderthal DNA in BK1653 and the IUP Bacho Kiro Cave individuals was more similar to the Vindija33.1910 and Chagyrskaya842 Neanderthals than to the Altai Neanderthal9 (2.8 ≤ |Z| ≤ 5.1; Supplementary Information 7). |
|
|
|
Post by Admin on May 3, 2022 21:32:25 GMT
To study the spatial distribution of Neanderthal ancestry in the genomes of the Bacho Kiro Cave individuals, we used around 1.7 million SNPs at which Neanderthal9 and/or Denisovan43 genomes differ from African genomes7 and an approach44 that detects tracts of archaic DNA in ancient genomes. We found a total of 279.6 centiMorgans (cM) of Neanderthal DNA in F6-620, 251.6 cM in CC7-335 and 220.9 cM in BB7-240, and these individuals carried seven, six and nine Neanderthal DNA segments longer than 5 cM, respectively (Fig. 3, Extended Data Fig. 7a, Supplementary Information 8). The longest introgressed Neanderthal segment in F6-620 encompassed 54.3 cM, and the longest segments in CC7-335 and BB7-240 were 25.6 cM and 17.4 cM, respectively (Fig. 3, Extended Data Fig. 7a). By contrast, the total amount of Neanderthal DNA in the BK1653 genome was 121.7 cM and the longest Neanderthal segment was 2.5 cM (Fig. 3, Extended Data Fig. 7a). Fig. 3: Geographical distribution of Neanderthal archaeological sites and genome-wide distribution of Neanderthal alleles in the genomes of ancient modern humans. 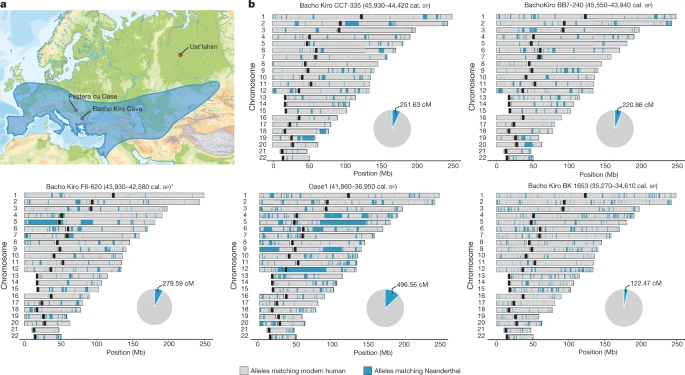 a, Neanderthal geographical range (blue) and the locations of Peştera cu Oase, Bacho Kiro Cave and where the femur of the Ust’Ishim individual was found. b, Distribution of Neanderthal DNA in ancient modern human genomes. Neanderthal DNA segments longer than 0.2 cM are indicated in blue. Pie charts indicate the total proportion of Neanderthal DNA identified in each genome. Centromeres are shown in black. On the basis of the distribution of the long Neanderthal segments, we estimate that individual F6-620 had a Neanderthal ancestor less than six generations back in his family tree (Extended Data Table 2, Supplementary Information 8). Both the CC7-335 and BB7-240 individuals had Neanderthal ancestors about seven generations back in their families, with upper confidence intervals of ten and seventeen generations, respectively (Extended Data Table 2, Extended Data Fig. 7b, Supplementary Information 8). Thus, all IUP Bacho Kiro Cave individuals had recent Neanderthal ancestors in their immediate family histories. To further explore the extent to which the Bacho Kiro Cave individuals contributed ancestry to later populations in Eurasia, we investigated whether the Neanderthal DNA segments in Bacho Kiro Cave genomes overlapped with Neanderthal segments in present-day populations more than expected by chance. We found more overlapping of segments between present-day East Asian populations and the IUP Bacho Kiro Cave individuals (lowest correlation coefficient of 0.09, 95% CI: 0.08–0.1) than with the BK1653 individual (P = 0.02, Wilcoxon test). By contrast, the BK1653 individual shows more overlapping of Neanderthal segments with present-day western Eurasian populations (a correlation coefficient of 0.11, 95% CI: 0.1–0.12) than do the IUP Bacho Kiro Cave individuals (P < 1 × 10−18, Wilcoxon test). This is compatible with the observation that the IUP Bacho Kiro Cave population contributed more ancestry to later populations in Asia and the Americas, whereas the BK1653 individual contributed more ancestry to populations in western Eurasia. We next looked for overlap between parts of the human genome that carry little or no Neanderthal ancestry (Neanderthal ‘deserts’), which are thought to have been caused by purifying selection against Neanderthal DNA shortly after introgression45,46. We find almost no introgressed Neanderthal DNA in the previously described deserts in the IUP Bacho Kiro Cave and Oase1 individuals (249 kb out of 898 Mb of introgressed sequence; P = 0.0079, permutation P value). When we restricted these comparisons to the more recent Neanderthal contributions (that is, segments longer than 5 cM), we similarly found no overlap (0 Mb out of 415 Mb, P = 0.15, permutation P value), suggesting that selection against Neanderthal DNA variants occurred within a few generations, although additional individuals with recent Neanderthal ancestry will be needed to fully resolve this question. In conclusion, the Bacho Kiro Cave genomes show that several distinct modern human populations existed during the early Upper Palaeolithic in Eurasia. Some of these populations, represented by the Oase1 and Ust’Ishim individuals, show no detectable affinities to later populations, whereas groups related to the IUP Bacho Kiro Cave individuals contributed to later populations with Asian ancestry as well as some western Eurasian humans such as the GoyetQ116-1 individual in Belgium. This is consistent with the fact that IUP archaeological assemblages are found from central and eastern Europe to present-day Mongolia5,15,16 (Fig. 1), and a putative IUP dispersal that reached from eastern Europe to East Asia. Eventually populations related to the IUP Bacho Kiro Cave individuals disappeared in western Eurasia without leaving a detectable genetic contribution to later populations, as indicated by the fact that later individuals, including BK1653 at Bacho Kiro Cave, were closer to present-day European populations than to present-day Asian populations29,30. In Europe, the notion of successive population replacements is also consistent with the archaeological record, where the IUP is clearly intrusive against the Middle Palaeolithic background and where, apart from the common focus on blades, there are no clear technological connections between the IUP and the subsequent Aurignacian technologies. Finally, it is striking that all four of the European individuals who overlapped in time with late Neanderthals7 and from whom genome-wide data have been retrieved had close Neanderthal relatives in their family histories (Fig. 3, Extended Data Figs. 7, 8). This suggests that mixing between Neanderthals and the first modern humans that arrived into Europe was perhaps more common than is often assumed. |
|

