|
|
Post by Admin on Sept 22, 2015 13:42:22 GMT
 Adolf Hitler is likely to have been descended from both Jews and Africans, according to DNA tests. Hitler's DNA was found to contain Haplogroup E1b1b, commonly found in the Berbers of Morocco and also accounts for approximately 18 to 20 per cent of the Y-chromosomes of Sephardic Jews who hail from Morocco, Spain and Portugal. A chromosome called Haplopgroup E1b1b (Y-DNA) in their samples is rare in Germany and indeed Western Europe. 'It is most commonly found in the Berbers of Morocco, in Algeria, Libya and Tunisia as well as among Ashkenazi and Sephardic Jews,' Mr Vermeeren said. 'This is a surprising result,' said Ronny Decorte, a genetic specialist who agreed that Hitler probably did have some roots in North Africa. E-M215 has two ancient branches that contain all known modern E-M215 men, E-M35 and E-M281. Of these two, the only branch that has been confirmed in a native population outside of Ethiopia is E-M35, which in turn has two known branches, E-V68, E-Z827 which contain by far the majority of all modern E-M215 men. E-V68 and E-V257 have been found in highest numbers in North Africa and the Horn of Africa; but also in lower numbers in parts of the Middle East and Europe, and in isolated populations of Southern Africa. |
|
|
|
Post by Admin on Oct 16, 2015 14:24:42 GMT
Researchers believe that our species evolved in East Africa around 190,000 to 160,000 years ago. After this, human fossils from the archaeological sites of Es Skhül and Jabel Qafzen, in Israel, suggest they spread there between 80,000 and 120,000 years ago. However, these samples preserve some archaic features that place them “on verge of modernity” but not fully modern yet. They have therefore been thought of as the result of a failed dispersal out of Africa that barely managed to reach beyond the borders of the African continent. So that means that modern humans were at the gates of Europe, but it took them another 50,000 years to be able to enter the land of the Neanderthal.  Genetic studies have indicated that the earliest successful “out of Africa” migration was around 60,000 years ago. But the newly discovered teeth, first found by a team of researchers at the Institute of Vertebrate Paleontology and Paleoanthropology of Beijing in the Fuyan (Daoxian) cave in southern China, are truly modern. The team dated the sample to be more than 80,000 years old, indicating that our species was present in Asia considerably earlier than had previously been suspected. In fact, it implied that anatomically fully modern humans lived in Asia 30,000 to 70,000 years earlier than in Europe and the eastern Mediterranean. The team’s estimation that the teeth could be as old as 120,000 years was hard to believe at first. That was a big claim to make. So when they invited us to come and visit the site, we were eager to go. In this case, the challenge was not to prove the species the teeth belonged to, but to be sure about the context and the dating of the teeth.  The teeth were found in a layer that was sealed by a continuous calcite floor, like an enormous gravestone that would have made it impossible for any soil or more recent fossils to accumulate below. There was a small stalagmite, an upward-growing pillar of mineral deposits from water dripping in, on top of this flowstone that experts from China and US have dated to be around 80,000 years old. As the stalagmite was formed after the calcite floor sealed the layer with the fossils, everything below had to be older than that.  The human teeth were consistently mixed with mammalian remains, including extinct hyenas, pandas and elephants that helped to infer a maximum age of 120,000 for the human fossils. The context was clear and we spent hours inside the cave concluding that there were no doubt that the evidence were consistently pointing towards the beginning of the Late Pleistocene age. From that moment, we remember the rest of our trip in China with the dizziness of the excitement and the shock. But also the warm welcome of the local people from Fuyan who celebrated our visit with all sort of luxury food and drinks. However, it was us who should pay the honour of having visited the earliest known site in the world with fully modern humans outside Africa. 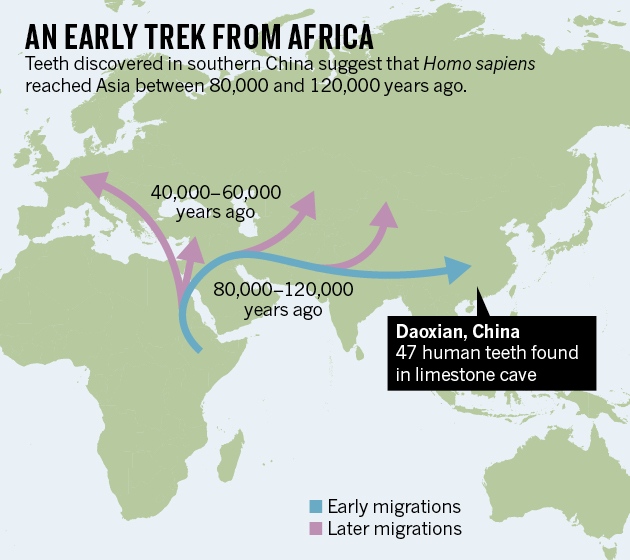 Those ages buck the conventional wisdom that H. sapiens from Africa began colonizing the world only around 50,000–60,000 years ago, says Martinón-Torres. Older traces of modern humans have been seen outside Africa, such as the roughly 100,000-year-old remains from the Skhul and Qafzeh Caves in Israel. But many researchers had argued that those remains were only evidence of unsuccessful efforts at wider migration. “This demonstrates it was not a failed dispersal,” says Petraglia, who has long argued for an early expansion of modern humans through Asia on a southerly route. “This is a rock-solid case for having early humans — definitely Homo sapiens — at an early date in eastern Asia.” Chris Stringer, a palaeoanthropologist at the Natural History Museum in London who had argued that remains from Skhul and Qafzeh signified unsuccesful migrations, says that he is now swayed by the Daoxian teeth.  It is also not clear why modern humans would have reached East Asia so long before they reached Europe, where the earliest remains are about 45,000 years old. Martinón-Torres suggests that humans could not gain a foothold in Europe until Neanderthals there were teetering on extinction. The frigid climate of Ice Age Europe may have erected another barrier to people adapted to Africa, says Petraglia. Although Hublin says there is a good case that the Daoxian teeth are older than 80,000 years, he notes that several of the teeth have visible cavities, a feature uncommon in human teeth older than 50,000 years. “It could be that early modern humans had a peculiar diet in tropical Asia,” he says. “But I am pretty sure that this observation will raise some eyebrows." Martinon-Torres says her team plans to look more closely at the cavities and the diet of the Daoxian humans by examining patterns of tooth wear. Nature doi:10.1038/nature.2015.18566 |
|
|
|
Post by Admin on Oct 27, 2015 13:55:01 GMT
 Figure 1: PCA of high-density array data. The EIGENSOFT package24 was used to identify the principal components (PCs) of autosomal variation in our Old World sample set (Fig. 1 and Supplementary Fig. 2a). This analysis places the studied samples along two well-established geographic axes of global genetic variation18,19,22: PC1 (sub-Saharan Africa versus the rest of the Old World) and PC2 (east versus west Eurasia). Focusing on the Middle Eastern populations in the PC1–PC2 plot (Fig. 1b) reveals more geographically refined groupings. Populations of the Caucasus, flanked by Cypriots, form an almost uninterrupted rim that separates the bulk of Europeans from Middle Eastern populations. Bedouins, Jordanians, Palestinians and Saudi Arabians are located in close proximity to each other, which is consistent with a common origin in the Arabian Peninsula25, whereas the Egyptian, Moroccan, Mozabite Berber, and Yemenite samples are located closer to sub- Saharan populations (Fig. 1a and Supplementary Fig. 2a). Most Jewish samples, other than those from Ethiopia and India, overlie non-Jewish samples from the Levant (Fig. 1b). The tight cluster comprising the Ashkenazi, Caucasus (Azerbaijani and Georgian), Middle Eastern (Iranian and Iraqi), north African (Moroccan) and Sephardi (Bulgarian and Turkish) Jewish communities, as well as Samaritans, strongly overlaps Israeli Druze and is centrally located on the principal component analysis (PCA) plot when compared with Middle Eastern, European Mediterranean, Anatolian and Caucasus non-Jewish populations (Fig. 1).  Figure 2: PCA of west Eurasian high-density array data. This Jewish cluster consists of Contemporary Jews comprise an aggregate of ethno-religious communities whose worldwide members identify with each other through various shared religious, historical and cultural traditions1,2. Historical evidence suggests common origins in the Middle East, followed by migrations leading to the establishment of communities of Jews in Europe, Africa and Asia, in what is termed the Jewish Diaspora3–5. This complex demographic history imposes special challenges in attempting to address the genetic structure of the Jewish people6. Although many genetic studies have shed light on Jewish origins and on diseases prevalent among Jewish communities, including studies focusing on uniparentally and biparentally inherited markers7–16, genome-wide patterns of variation across the vast geographic span of Jewish Diaspora communities and their respective neighbours have yet to be addressed. Here we use high-density bead arrays to genotype individuals from 14 Jewish Diaspora communities and compare these patterns of genome-wide diversity with those from 69 Old World non-Jewish populations, of which 25 have not previously been reported. These samples were carefully chosen to provide comprehensive comparisons between Jewish and non-Jewish populations in the Diaspora, as well as with non-Jewish populations from the Middle East and north Africa. Principal component and structure-like analyses identify previously unrecognized genetic substructure within the Middle East. Most Jewish samples form a remarkably tight subcluster that overlies Druze and Cypriot samples but not samples from other Levantine populations or paired Diaspora host populations.  Figure 3: Population structure inferred by ADMIXTURE analysis. In contrast, Ethiopian Jews (Beta Israel) and Indian Jews (Bene Israel and Cochini) cluster with neighbouring autochthonous populations in Ethiopia and western India, respectively, despite a clear paternal link between the Bene Israel and the Levant. These results cast light on the variegated genetic architecture of the Middle East, and trace the origins of most Jewish Diaspora communities to the Levant. Recently, the capacity to obtain whole-genome genotypes with the use of array technology has provided a robust tool for elucidating fine- scale population structure and aspects of demographic history17–23. This approach, initially used to account for population stratification in genome-wide association studies, identified genome-wide patterns of variation that distinguished between Ashkenazi Jews and non- Jews of European descent7,11,12,14–16. Similarly, a large-scale survey of autosomal microsatellites found that samples from four Jewish communities clustered close to each other and intermediate between non-Jewish Middle Eastern and European populations10. Illumina 610K and 660K bead arrays were used to genotype 121 samples from 14 Jewish communities. The results were compared with 1,166 individuals from 69 non-Jewish populations (Supplementary Note 1 and Supplementary Table 1), with particular attention to neighbouring or ‘host’ populations in corresponding geographic regions. These results were also integrated with analyses of genotype data from about 8,000 Ychromosomes and 14,000 mitochondrial DNA (mtDNA) samples (Supplementary Note 6 and Supplementary Tables 4 and 5).  Although uniparental markers8,9 (Supplementary Note 6) are limited in their capacity to uncover genetic substructure within the Middle East, they do provide important insights into sex-specific processes that are not unambiguously evident from the autosomal data alone. For example, Y-chromosome data point to a unique paternal genetic link between the Bene Israel community and the Levant, whereas the absence of sub-Saharan African maternal lineages in Yemenite and Moroccan Jews (in contrast to their hosts) suggests limited maternal gene flow. Nature 466, 238–242 (08 July 2010) doi:10.1038/nature09103 |
|
|
|
Post by Admin on Dec 7, 2015 7:43:38 GMT
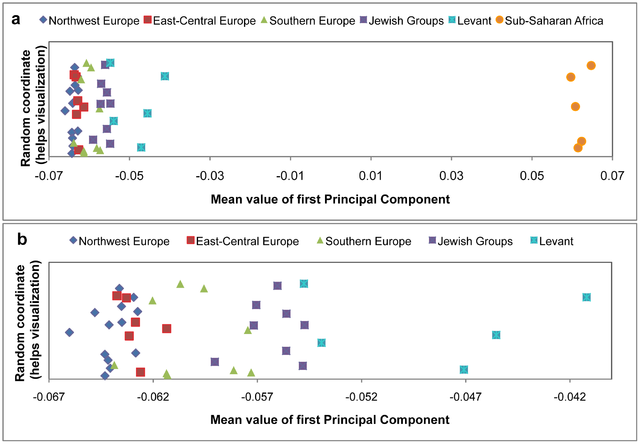 Figure 1. PCA Projection. The history of human migrations from Africa into West Eurasia is only partially understood. Archaeological and genetic evidence indicate that anatomically modern humans arrived in Europe from an African source at least 45,000 years ago, following the initial dispersal out of Africa [1], [2]. However, it is known that Southern Europeans and Levantines (people from modern day Palestine, Israel, Syria and Jordan) have also inherited genetic material of African origin due to subsequent migrations. One line of evidence comes from Y-chromosome [3] and mitochondrial DNA analyses [4]–[6]. These have identified haplogroups that are characteristic of sub-Saharan Africans in Southern Europeans and Levantines but not in Northern Europeans [7]. Auton et al. [8] presented nuclear genome-based evidence for sharing of sub-Saharan African ancestry in some West Eurasians, by identifying a North-South gradient of haplotype sharing between Europeans and sub-Saharan Africans, with the highest proportion of haplotype sharing observed in south/southwestern Europe. However, none of these studies used genome-wide data to estimate the proportion of African ancestry in West Eurasians, or the date(s) of mixture. Throughout this report, we use “African mixture” to refer to gene flow into West Eurasians since the divergence of the latter from East Asians; thus, we are not referring to the much older dispersal out of Africa ∼45,000 years ago but instead to migrations that have occurred since that time. To study the signal of African gene flow into West Eurasian populations, we began by computing principal components (PCs) using San Bushmen (HGDP-CEPH- San) and East Eurasians (HapMap3 Han Chinese- CHB), and plotted the mean values of the samples from each West Eurasian population onto the first PC, a procedure called “PCA projection” [17], [18]. The choice of San and CHB, which are both diverged from the West Eurasian ancestral populations [19], [20], ensures that the patterns in PCA are not affected by genetic drift in West Eurasians that has occurred since their common divergence from East Eurasians and South Africans. We observe that many Levantine, Southern European and Jewish populations are shifted towards San compared to Northern Europeans, consistent with African mixture, and motivating formal testing for the presence of African ancestry (Figure 1, Figure S2). 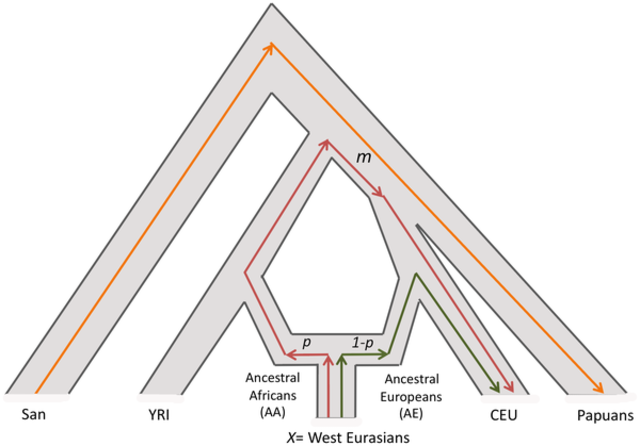 Figure 2. Estimation of African ancestry using f4 Ancestry Estimation. To estimate the proportion of sub-Saharan African ancestry in the various West Eurasian populations that showed significant evidence of mixture, we used f4 Ancestry Estimation [21], a method which produces accurate estimates of ancestry proportions, even in the absence of data from the true ancestral populations. This method estimates mixture proportions by fitting a model of mixture between two ancestral populations, followed by (possibly large) population-specific genetic drift. Briefly, we calculate a statistic that is proportional to the correlation in the allele frequency difference between West Eurasians and sub-Saharan Africans, and divide it by the same statistic for a population of sub-Saharan African ancestry, like YRI (Figure 2). This method has been shown through simulation to be robust to ascertainment bias on the SNP arrays and deviations from the assumed model of mixture (e.g. date and number of mixture events) [21]. The finding of sub-Saharan African ancestry in West Eurasians predicts that there will be a signature of admixture LD in the populations that experienced this mixture. That is, there will be LD between all markers that are highly differentiated between the two ancestral populations and the allele will be strongly correlated to the local ancestry [23]. Hence, there will be chromosomal segments of African ancestry with lengths that reflect the number of recombination events that have occurred since mixture, and thus can be used to estimate an admixture date. Figure 3 shows that this expected pattern is observed empirically in the decay of LD in four example West Eurasian populations, where we enhance the effects of admixture LD by weighting the SNP comparisons by frequency difference between the ancestral Africans (YRI) and ancestral West Eurasians (CEU). In the Southern European, Jewish and Levantine populations, this procedure produces clear evidence of admixture LD (Figure 3). However, Northern Europeans (Russians in Figure 3) do not show any evidence of African gene flow, consistent with the 4 Population and 3 Population Test results and Figure 1. Similar results are seen for other West Eurasian and Jewish populations that show evidence of mixture in the 4 Population Test. 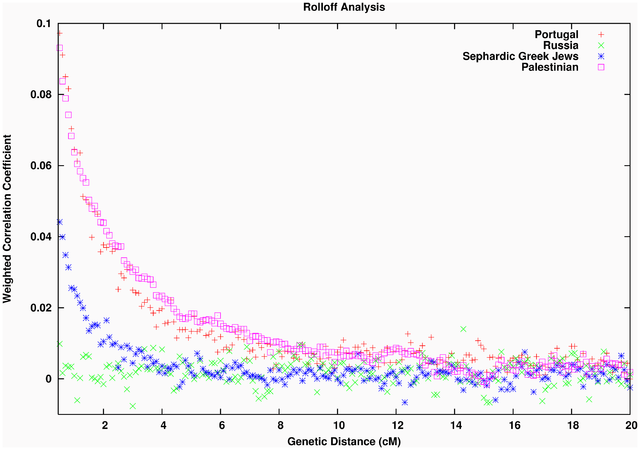 Figure 3. Testing for LD due to African admixture in West Eurasians. The finding of African ancestry in Southern Europe dating to ∼55 generations ago, or ∼1,600 years ago assuming 29 years per generation [30], needs to be placed in historical context. The historical record documents multiple interactions of African and European populations over this period. One potential opportunity for African gene flow was during the period of Roman occupation of North Africa that lasted until the early 5th century AD, and indeed tomb inscriptions and literary references suggest that trade relations continued even after that time [31], [32]. North Africa was also a supplier of goods and products such as wine and olive oil to Italy, Spain and Gaul from 200–600 AD, and Morocco was a major manufacturer of the processed fish sauce condiment, garum, which was imported by Romans [33]. In addition, there was slave trading across the western Sahara during Roman times [7], [34]. Another potential source of some of the African ancestry, especially in Spain and Portugal, is the invasion of Iberia by Moorish armies after 711 AD [35], [36]. If the Moors already had some African ancestry when they arrived in Southern Europe, and then admixed with Iberians, we would expect the admixture date to be older than the date of the invasion, as we observe. The signal of African mixture that we detect in Levantines (Bedouins, Palestinians and Druze) – an average of 32 generations or ∼1000 years ago – is more recent than the signal in Europeans, which might be related to the migrations between North Africa and Middle East that have occurred over the last thousand years, and the proximity of Levantine groups geographically to Africa. Syria and Palestine were under Egyptian political control until the 16th century AD when they were conquered by the Ottoman Empire. This is in concordance with our proposed dates. In addition, the Arab slave trade is responsible for the movement of large numbers of people from Africa across the Red Sea to Arabia from 650 to 1900 AD and probably even prior to the Islamic times [7], [37]. We caution that our sampling of the Middle East is sparse, and it will be of interest to study African ancestry in additional groups from this region. A striking finding from our study is the consistent detection of 3–5% sub-Saharan African ancestry in the 8 diverse Jewish groups we studied, Ashkenazis (from northern Europe), Sephardis (from Italy, Turkey and Greece), and Mizrahis (from Syria, Iran and Iraq). This pattern has not been detected in previous analyses of mitochondrial DNA and Y chromosome data [7], and although it can be seen when re-examining published results of STRUCTURE-like analyses of autosomal data, it was not highlighted in those studies, or shown to unambiguously reflect sub-Saharan African admixture [15], [38]. We estimate that the average date of the mixture of 72 generations (∼2,000 years assuming 29 years per generation [30]) is older than that in Southern Europeans or other Levantines. The point estimates over all 8 populations are between 1,600–3,400 years ago, but with largely overlapping confidence intervals. It is intriguing that the Mizrahi Irani and Iraqi Jews—who are thought to descend at least in part from Jews who were exiled to Babylon about 2,600 years ago [39], [40]—share the signal of African admixture. (An important caveat is that there is significant heterogeneity in the dates of African mixture in various Jewish populations.) A parsimonious explanation for these observations is that they reflect a history in which many of the Jewish groups descend from a common ancestral population which was itself admixed with Africans, prior to the beginning of the Jewish diaspora that occurred in 8th to 6th century BC [41]. The dates that emerge from our ROLLOFF analysis in the non-Mizrahi Jews could also reflect events in the Greek and Roman periods, when there were large communities of Jews in North Africa, particularly Alexandria [34], [42]. We detect a similar African mixture proportion in the non-Jewish Druze (4.4±0.4%) although the date is more recent (54±7 generations; 44±7 after the bias correction). Algorithms such as PCA and STRUCTURE show that various Jewish populations cluster with Druze [15], which coupled with the similarity in mixture proportions, is consistent with descent from a common ancestral population. Importantly, the other Levantine populations (Bedouins and Palestinians) do not share this similarity in the African mixture pattern with Jews and Druze, making them distinct in their admixture history. Moorjani P, Patterson N, Hirschhorn JN, Keinan A, Hao L, Atzmon G, et al. (2011) The History of African Gene Flow into Southern Europeans, Levantines, and Jews. PLoS Genet 7(4): e1001373. doi:10.1371/journal.pgen.1001373 |
|
|
|
Post by Admin on Dec 31, 2015 7:18:52 GMT
 As historians of the Soviet multiethnic state have argued, the Soviet state itself was a “maker of nations,” institutionalizing ethnicity by creating territorial republics based on ethnic criteria and promoting “national languages” and "national cultures" within them. The state also practiced a kind of affirmative action, actively recruiting members of underrepresented nationalities into higher education, jobs, and Communist Party membership.[2] Every Soviet citizen had a single nationality, fixed at the age of 16 and registered in his or her identity document. Being “mixed,” in this context, was not understood as interracial (as in the United States) or interfaith (as in pre-revolutionary tsarist Russia); it was “interethnic” or sometimes “inter-national.”  Contrasting sharply with the anti-miscegenation attitudes prevalent in the United States, the Soviet state beginning in the early 1930s supported ethnic mixing both in theory and practice. This was originally a response to the eugenicist ideas then prevalent in Nazi Germany and elsewhere. Soviet theorists attacked their ideological opponents by arguing that differences between groups were due to history and culture, not biology. All ethnic groups were equally capable of flourishing, given the right (i.e. socialist) conditions. Moreover, Soviet anthropologists challenged the notion that ethnic or “racial” mixing led to degeneracy and pathology—an idea that had wide currency in the 1930s not just in Germany, but among West European and American scholars. Soviet anthropologists conducted studies designed to show that mixed individuals were just as healthy and productive as people of supposedly “pure” racial background. Ethnic mixing, in the official view, was a positive manifestation of social progress.[3]  In the world’s first socialist society, intermarriage was seen as contributing to the eventual merging of the Soviet nations into a single “Soviet people.” Intermarriage was also closely associated with modernity, and in particular with the arrival of modernity in “backward” areas such as Central Asia. Soviet analysts argued that mixed couples were more likely to abandon traditional ways and adopt a modern, typically Soviet lifestyle, thereby setting an example for other Central Asians to follow. This “modernity” was often understood in terms of Russian-ness or European-ness, since mixed families were more likely to speak Russian at home and lead an urban, European lifestyle. (Although there was no official policy of russification in the USSR, Russian was the dominant culture and non-Russians were encouraged to become competent in Russian as a second language.) Official Soviet policy had certain similarities with the Latin American ideology of mestizaje, which also celebrated multiethnic and multiracial hybridity.[4] The Soviet approach to intermarriage in Central Asia, with its overtones of Russification, also resembles the attempts of Australia and New Zealand to “Europeanize” their indigenous populations through ethnic mixing (though Russification in the Soviet case was always envisioned as cultural and not biological).[5] |
|

