|
|
Post by Admin on Jan 7, 2023 18:21:51 GMT
 Fig. 3. Graph modelling of present-day Apulians and Greeks. Z scores of both trees are between -3 and 3. Ancestries highlighted in shades of red are ancestries specifically shared with Bronze Age Greeks. Letters from A to F refer to unsampled ancestral populations. A) modelling of Apulians, B) modelling of Greek population. 2.3. Investigating putative signature of adaptation among Northern and Southern Italians Following the differences in ancient ancestry contributions across the Italian Peninsula, we applied the Population Branch Statistics (PBS) analysis in order to identify markers showing significant allele frequency differences among pairs of Italian populations using CHB as an outgroup. We estimated the average PBS across windows of five consecutive SNPs and focused on the top ten windows with higher frequency in Northern (positive PBS values) and Southern Italy (negative PBS values), respectively (Fig. S5). We finally annotated the variants with the most extreme PBS values (above the 99.5 percentile and below 0.05) and evaluated if they were enriched for any biological or functional feature. Among the top 20 windows, seven do not overlap with any known gene (Fig. 4, Table S3). Of those showing the highest allele frequencies in Northern Italy, one encompasses PABPC4L (H. sapiens poly(A) binding protein, cytoplasmic 4-like), a gene associated with venous thromboembolism [49], birth weight [50], blood protein level and alcohol consumption [51]; another, located on chromosome 1, encompasses the gene NID1 (Nidogen 1), which is associated with blood proteins [52] and platelet levels [53,54], and a third on chromosome 3 maps within the CBLB (Cbl proto-oncogene, E3 ubiquitin protein ligase B) gene, involved in the immune response by regulating T- and B- cell receptors [55]. |
|
|
|
Post by Admin on Jan 13, 2023 20:16:08 GMT
2.3. Investigating putative signature of adaptation among Northern and Southern Italians Following the differences in ancient ancestry contributions across the Italian Peninsula, we applied the Population Branch Statistics (PBS) analysis in order to identify markers showing significant allele frequency differences among pairs of Italian populations using CHB as an outgroup. We estimated the average PBS across windows of five consecutive SNPs and focused on the top ten windows with higher frequency in Northern (positive PBS values) and Southern Italy (negative PBS values), respectively (Fig. S5). We finally annotated the variants with the most extreme PBS values (above the 99.5 percentile and below 0.05) and evaluated if they were enriched for any biological or functional feature. Among the top 20 windows, seven do not overlap with any known gene (Fig. 4, Table S3). Of those showing the highest allele frequencies in Northern Italy, one encompasses PABPC4L (H. sapiens poly(A) binding protein, cytoplasmic 4-like), a gene associated with venous thromboembolism [49], birth weight [50], blood protein level and alcohol consumption [51]; another, located on chromosome 1, encompasses the gene NID1 (Nidogen 1), which is associated with blood proteins [52] and platelet levels [53,54], and a third on chromosome 3 maps within the CBLB (Cbl proto-oncogene, E3 ubiquitin protein ligase B) gene, involved in the immune response by regulating T- and B- cell receptors [55].  Fig. 4. Karyotype plot of the top 10 windows from PBS analysis. In red and blue are indicated the chromosome locations for the top 10 PBS in North and South Italy, respectively. Where present, the set of annotated genes is presented. Of the windows with extreme negative PBS values, the two most divergent are located within chromosome 12 and encompass 15 genes (Fig. 4, Table S3). They include ALDH2, known to affect the metabolism of alcohol and correlated to different behavioural traits associated with alcohol consumption [56]. We note here that our dataset does not contain SNPs within this gene, and it is thus possible that this signal is led by variants in other regions. Finally, we found extreme divergence in a region within the EDAR (Ectodysplasin A receptor) gene, associated with variations in head hair morphology and facial hair thickness [57]. Although the SNP rs3827760 (chr2:109513601), affecting hair follicle thickness and shape, is not present in our dataset, it is unlikely that it is driving the adaptation signal, as also observed in a previous exome analysis [38]. 3. Discussion Since prehistoric times Southern Italy has been a cross-roads of many human migrations and interactions, and their genetic signatures are still present in the DNA of individuals currently living in this area [17,41,58,59]. In this study, we highlighted a high similarity between Southern Italy and the Peloponnese. In fact, our cluster analysis showed that present-day South-Eastern Peloponnesian populations have high genetic affinity with modern Apulians, Calabrians and South-Eastern Sicilians, all characterised by a cluster composition different from those displayed by other Greek groups (Fig. 1B, Fig. S3). Additionally, individuals from Western Sicily show similarities with populations inhabiting the Western part of Peloponnese (Fig. 1B, Fig. S4). Although establishing the chronological context for this affinity using present-day genomes might be challenging, our results are in accordance with archaeological and historical sources that attributed the origin of Greek colonies in South-Eastern Sicily and Apulia from populations inhabiting the southern and Eastern parts of the Peloponnese [31,32]. Uniparental Y-chromosome findings are also in agreement with these observations revealing Eastern Peloponnesian ancestries in East Sicily [33] and shared haplogroups among modern-day Greeks and populations living in Southern Italian areas colonised by Greeks such as the Salento (Apulia) and the Ionian coast of Calabria [60]. The lower affinity with other Balkan populations could be attributed to a lower influence by inland populations, such as Slavic-related people [61] and/or genetic drift in Tsakones and Maniots as suggested by historical sources [39]. However the observation that, in some analyses, Southern Italians and ancient Greeks share more alleles than modern and ancient Peloponnesians, may suggest a scenario including the preservation of an ancient population signal in the genome of Southern Italians that was likely diluted by inland migrations in Greece (Fig. 3). Therefore, our results imply a high affinity between Southern Italians and Peloponnesians, with some differentiation among the two sides of the Adriatic Sea possibly due to recent demographic events such as migrations and/or admixture as the one recorded during the Middle Age period [62]. Notably, a substantial genetic affinity was highlighted between Peloponnesian individuals from the Neolithic and the Bronze Age, whose signatures are also recorded in Italy. Recent genomic studies [[17], [34], [37]] detected a contribution ultimately derived from the Caucasus (CHG) in modern Southern Italians and likely brought to Italy as early as the Bronze Age, although their demographic dynamics are still unknown. Overall, these results are in agreement with the detection of a small proportion of Iranian-related ancestry in Sicilian Middle Bronze Age samples [18], which could be tentatively linked to the spread of the Mycenaean culture [63]. Interestingly, our results modelled the source of this contribution as a mixture of AN and Iran Neolithic ancestries [14]. The latter was found consistently across Southern Italy and the Peloponnese, confirming similar again common genetic sources ancestral profiles shared between these two regions (Fig. 2C, D). Furthermore, our comparison between ancient and modern samples, showing an overall similarity between Southern Italian and Iron Age Individuals, may suggest that the CHG/Iran_Neolithic signature reached the East side of the Adriatic Sea multiple times, or possibly as a continuous gene flow. The largest degree of genetic heterogeneity across the European continent has been so far recorded in the Italian Peninsula [17], an important aspect to consider when epidemiological and translational studies in Italy are planned. Recent researches taking advantage of high coverage whole-genome and whole-exome sequences linked variants under selection in Italians to genes that were related to insulin secretion, obesity, thermogenesis, alcohol consumption, pathogenic-response, skin colour and cancer [37,38]. Here we assessed the occurrence of extreme differences in allele frequencies between Northern and Southern Italy close to the ALDH2, NID1, and CBLB genes that may have a role in alcohol metabolism, nevi features and immunological traits, respectively. Although the analysed dataset contained only ~ 100k SNPs, this replication in the possibly affected phenotypes may suggest an actual important difference among groups on the two parts of the peninsula. Nevertheless, genomic scans are sometimes controversial and difficult to interpret, and a deeper investigation, possibly involving multiple tests on denser datasets, is highly desirable. In conclusion, we provided new insights on the composition of modern genomes of individuals collected from Southern Italy, a still poorly sampled region of Europe. The fine characterisation of the time of arrival and routes of migrations related to the detected ancient signals is expected to be addressed only by the analysis of Apulian ancient genomes covering a period ranging from the Late Neolithic to the Early and Middle Bronze Age periods. The differences identified in modern-day Southern and Northern Italians have significant phenotypic implications and call for more extensive investigations on a larger number of markers and individuals. |
|
|
|
Post by Admin on Jan 16, 2023 17:28:21 GMT
Ancient DNA reveals admixture history and endogamy in the prehistoric Aegean
Abstract
The Neolithic and Bronze Ages were highly transformative periods for the genetic history of Europe but for the Aegean—a region fundamental to Europe’s prehistory—the biological dimensions of cultural transitions have been elucidated only to a limited extent so far. We have analysed newly generated genome-wide data from 102 ancient individuals from Crete, the Greek mainland and the Aegean Islands, spanning from the Neolithic to the Iron Age. We found that the early farmers from Crete shared the same ancestry as other contemporaneous Neolithic Aegeans. In contrast, the end of the Neolithic period and the following Early Bronze Age were marked by ‘eastern’ gene flow, which was predominantly of Anatolian origin in Crete. Confirming previous findings for additional Central/Eastern European ancestry in the Greek mainland by the Middle Bronze Age, we additionally show that such genetic signatures appeared in Crete gradually from the seventeenth to twelfth centuries BC, a period when the influence of the mainland over the island intensified. Biological and cultural connectedness within the Aegean is also supported by the finding of consanguineous endogamy practiced at high frequencies, unprecedented in the global ancient DNA record. Our results highlight the potential of archaeogenomic approaches in the Aegean for unravelling the interplay of genetic admixture, marital and other cultural practices.
Main
The Aegean has long been recognized as a region of major importance for understanding transregional societal transformations between Europe and the Near East. Already during the seventh millennium BC, the first farming communities emerged in the Aegean, whereby the earliest evidence was unearthed on the island of Crete—that is, the oldest occupation level below the later palace of Knossos1—but the origins of these populations remain ambiguous. The next major transformation in Aegean prehistory took place during the Early Bronze Age (EBA; about 3100–2000 BC). Complex societies emerged, characterized by sophisticated architecture, metallurgy, sealing systems and the integration of the Aegean in the Bronze Age Eastern Mediterranean networks of exchange. During the late third millennium BC, the Greek mainland witnessed a severe societal breakdown (at the end of Early Helladic II) with lasting impact until the later Middle Helladic period of the early second millennium2,3. This disruption has been attributed to various factors, among them dramatic climatic changes2,4,5 and the arrival of new groups6,7,8. Crete does not seem to have suffered a comparable period of decline9,10. With the emergence of the first palaces during the nineteenth century BC in the Middle Minoan period, the island’s societies transformed into a hitherto unknown sophistication in art, architecture and social practices.
Only a few centuries later, during the late Middle Bronze Age (MBA; Middle Helladic for the mainland), the first rich shaft graves of local elites appeared in southern mainland Greece, often displaying Minoan influences11. The competition between rising elites during the Shaft Grave period led to regional conflicts and culminated in the decline of many local dominions on the Greek mainland and possibly a first mainland military expedition to Crete during the fifteenth century12. At the end of this conflict, the palatial period (Late Helladic IIIA-B) started with a few eminent polities centred in Mycenae, Tiryns, Pylos, Athens, Hagios Vasileios in Laconia, Thebes, Orchomenos and Dimini—to name only the most prominent ones13,14,15. During this time, the influence on Crete by mainland centres intensified and Cretan resources were systematically exploited with the help of turning key palatial centres and cities like Knossos, Hagia Triada and Chania into outposts for the administration of large parts of the island16. So far, past human migrations in the Aegean were primarily reconstructed on the basis of archaeological and textual evidence but bioarchaeological studies have been adding new information during recent decades17,18,19,20,21,22.
Biomolecular approaches based on ancient DNA (aDNA) have been introduced in prehistoric Aegean research during the last decade. The first aDNA study analysed mitochondrial genomes23, emphasizing autochthonous developments rather than migration from outside Crete. Subsequent studies generated nuclear aDNA data and showed a common gene pool for the Aegean Neolithic populations, indicating that the southern Greek mainland differed from the northern in its higher genetic affinity to early Holocene populations from the Iran/Caucasus24,25. Others reported the presence of this ‘eastern’ (Iran/Caucasus-associated) genetic component in both Bronze Age (BA) Cretan (Minoan) and southern Greek mainland (Mycenaean) populations26. However, the last carried additional ancestry linked to the Western Eurasian Steppe herders (WES)27,28 or Armenia. Recently, Clemente and colleagues expanded the sampling scope of the BA Aegean to the northern mainland and the Aegean Islands corroborating the previous findings but also reporting higher WES-related ancestry in MBA individuals from northern Greece29.
Recent archaeogenetic studies outside the Aegean have engaged into integrating biological information as elements of the past social organization and structures30,31,32,33, whereby it is necessary to acknowledge that relational identities are not determined only through biological kinship34. Most approaches to past kinship in the Aegean were based on morphometric and non-metric analyses17,19,35 and first PCR-based studies were unsuccessful36. However, the potential of this line of evidence from the Aegean BA is outstanding due to the richness of collective burials as an expression and constitution of social belonging within local communities37.
|
|
|
|
Post by Admin on Jan 18, 2023 4:17:24 GMT
Results The archaeogenetic dataset Here, we generated new genome-wide data from 102 prehistoric individuals from Aegean Neolithic (n = 6), BA (n = 95), as well as Iron Age contexts (IA; n = 1) (Fig. 1 and Supplementary Note 1), thereby achieving a fourfold increase in sample size from previously published datasets. This sample, owing to the geographical and temporal distribution, enables us to address complex features of admixture history and other biological aspects interwoven into these prehistoric societies (for example, marital practices). Nea Styra on the island of Euboea and Lazarides on the island of Aegina add to the post-Neolithic sites included that date to the time before the debated disruption around 2200 BC (the end of Early Helladic II on the Greek mainland). The remaining individuals from the mainland and the islands are attributed to the Mycenaean culture of the Late Bronze Age (LBA) (Aidonia, Glyka Nera, Lazarides, Koukounaries, Mygdalia and Tiryns). Most of the data come from Crete (66 of 102 individuals), in a time transect that covers early phases of the Neolithic (Aposelemis; late seventh to early sixth millennia BC) and the BA (Hagios Charalambos—Early-Middle Minoan; Chania, Aposelemis and Krousonas—Late Minoan). With the exception of Aposelemis and XAN035 from Chania (about 1700–1450 BC), all other Late Minoan individuals date between about 1400 and 1100 BC (LMII–III). All the analysed skeletal remains from Nea Styra, Mygdalia, BA Aposelemis, Krousonas, Aidonia and Hagios Charalambos belonged to the same within-site collective burial context; for the latter, Yersinia pestis and Salmonella enterica were also recently detected38. Extracted aDNA was immortalized into genomic libraries, part of which were enriched for 1,233,013 ancestry-informative single nucleotide polymorphisms (SNPs) (1240K) (Methods) and sequencing data were evaluated for aDNA preservation and contamination (Supplementary Tables 1 and 2). In our inferences for the Aegean individuals, we re-appraised all previously published contemporaneous individuals from this area24,25,26,29 (Fig. 1). We also radiocarbon dated 43 of the skeletal remains that yielded genome-wide data (Supplementary Table 3; Methods). Fig. 1: Location and dates of individuals with newly generated aDNA data. 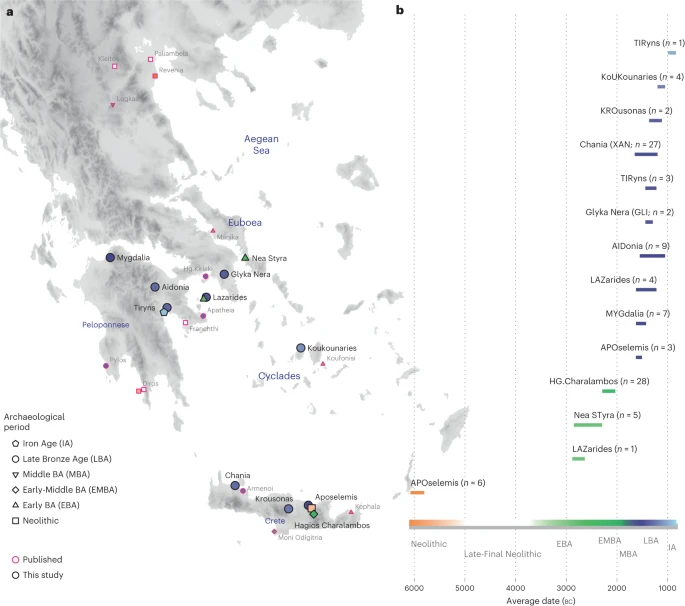 a, Geographical distribution of archaeological sites mentioned in the study annotated by period. Sites with smaller symbols of light outline refer to the published datasets that are co-analysed and follow the same symbol/colour scheme. Data obtained from the same site but different periods, are annotated with jittering points. b, The number of individuals analysed and their date range based on archaeological chronology or radiocarbon dating. Site names are abbreviated in three-letter capitalized identifiers as indicated in the labels. E, Early; M, Middle; L, Late. See also Supplementary Tables 2 and 3. Transregional genetic entanglements of Aegean populations To visualize genetic ancestry variation, we first performed a principal component analysis (PCA) on modern West Eurasian populations and projected onto the first two PCs the ancient individuals from the Aegean and nearby regions (Fig. 2). The six individuals from Neolithic Aposelemis cluster with other early European and Anatolian/Aegean farmers, suggesting that the gene pool of Neolithic Crete was linked to the broader Aegean during that period. After around two millennia, the EBA and MBA individuals show a substantial change in their PC coordinates, shifted along PC2 towards Early Holocene Iran/Caucasus and the descending Chalcolithic and BA Anatolians/ BA Caucasians. This shift does not seem uniform, as the five individuals from Nea Styra, who were buried together in the same shaft grave, show substantial genetic variation. Finally, the LBA individuals deviate from these earlier BA individuals towards BA Central and Eastern Europe, suggesting multiphased genetic shifts in the Aegean since the Neolithic. Fig. 2: West Eurasian PCA (grey background points) with projection of ancient Aegean and other ancient relevant populations (coloured points). 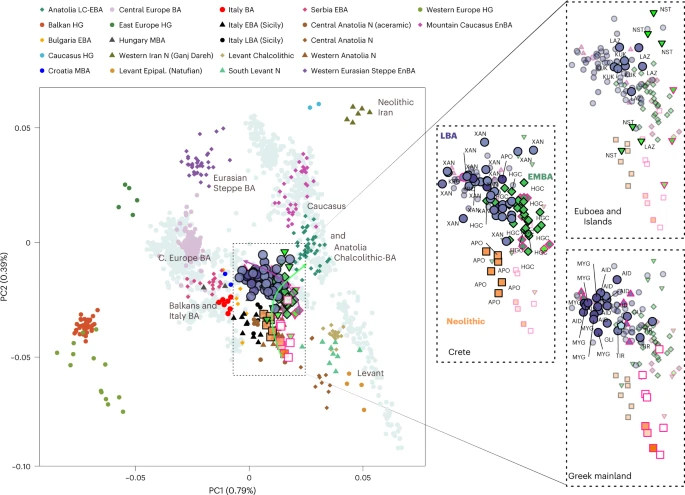 The arrows indicate the two major observed genetic shifts: from the Neolithic (N) to the EBA and from the MBA to the LBA. A zoom-in of coordinates for the Aegean samples is given and is subdivided by region (right). In every panel, the coordinates of the counterparts are plotted in the background in faded colours. The three-letter identifier of every individual is plotted as well. HG, hunter-gatherers; Epipal., Epipalaeolithic. To formally test whether the remarks from the PCA are consistent with diachronic gene-flow events, we used f-statistics of the form f4 (Mbuti, Test; Anatolian farmers, Aegean) (Methods; Supplementary Note 2) that contrast the various Aegean groups with the Anatolian farmers east of the Aegean (Supplementary Table 4). Affinities with far-eastern groups like Neolithic Iran are traced for Neolithic Aposelemis (or APO004) but only reach significance levels (≥3 s.e. or Z ≥ 3) on the EBA group from Nea Styra and then prevail for most of the later Aegean BA groups. However, the LBA ones additionally share alleles with contemporaneous or earlier (Mesolithic) populations from Central and Eastern Europe (for example, Eastern European hunter-gatherers: EEHG, Germany ‘Corded_Ware’, ‘Russia_Samara_EBA_Yamnaya’ and ‘Russia_North_Caucasus’). In addition, evidence of admixture from these groups was confirmed with admixture f3 test (Supplementary Table 5 and Supplementary Note 2). |
|
|
|
Post by Admin on Jan 19, 2023 5:35:25 GMT
Neolithic to Early/Middle Bronze Age Informed by the f-statistics, we explored formal admixture models using the software qpAdm (Methods; Supplementary Note 2). First, we tested a no-admixture model, which treated every individual as a sister group of Neolithic western Anatolia (‘W. Anatolia_N’) and then models by adding sequentially Neolithic Iran (‘W. Iran N’) and EEHG (Fig. 3). Substantial EEHG coefficients were fitted only on LBA and the two MBA individuals from the northern mainland ranging from around 5% to 25%, which explains why for some of them the simpler Anatolia + Iran Neolithic model was also adequate. Notably, Iran/Caucasus-related genetic influx was inferred in published individuals from the later Neolithic phases on the mainland (I2318, I709 and I3920; Peloponnese, around the fifth millenium BC)—but not earlier—as well as most of the EBA individuals from Euboea, Aegina and Koufonisia. Overall, the genetic heterogeneity among the Late Neolithic (LN) to EBA is not correlated with time alone, since within the Nea Styra grave male individuals carried substantially varying proportions of Iranian-related ancestry. By applying DATES on the LN and EBA individuals from the mainland and the islands (Methods), we obtained an average admixture date of around 4300 ± 250 BC (Supplementary Table 6), which is slightly younger when estimated from the Nea Styra individuals alone (about 3900 ± 460 BC). This variance in admixture dates also corroborates ongoing biological admixing with incoming individuals from the east of the Aegean following the establishment of the first Neolithic Aegean communities. Fig. 3: Admixture modelling with qpAdm per individual and group. 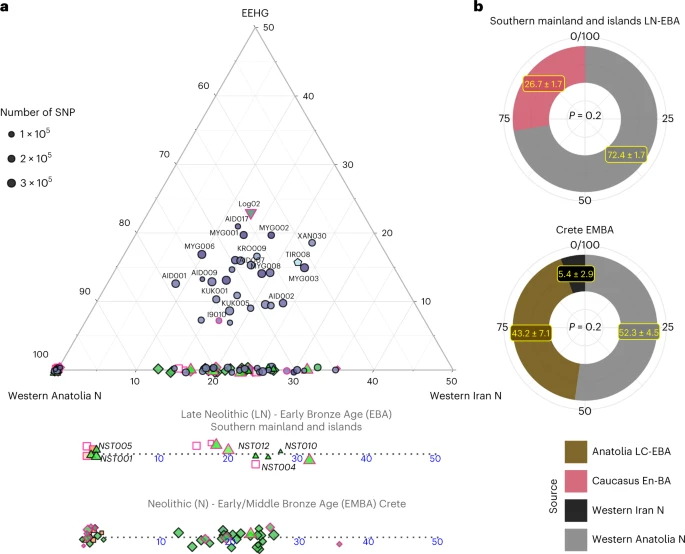 a, Ternary plot for a three-way admixture model of Aegean individuals using the distal sources of ceramic farmers from Western Anatolia, Western Iranian farmers from Ganj Dareh and the EEHG, all dating to about 6000 BC. Because qpAdm is based on allele frequency differences, modelling of individual targets has a lower resolution especially when the SNP coverage is low. A few of the Late-Final Neolithic (LN) and EBA individuals show additional ancestry related to Neolithic Western Iran. To better visualize the fluctuation or Iranian-like coefficients among the LN-E/MBA individuals, the Anatolian–Iranian axis is also provided separately for Crete and the mainland islands. Fitting models were chosen with a cutoff of P ≥ 0.01, with only four individuals falling in the lower range (0.01 ≤ P < 0.05). b, Allele frequencies are averaged among all LN-EBA individuals from the southern mainland and all EMBA Cretan individuals and modelled using proximal in time and space source populations. For the former, a source proxy from the Eneolithic/BA Caucasus fits better than Anatolia, whereas the opposite holds for Crete. However, for the latter, the model becomes adequate with the inclusion of additional low contribution from Neolithic Iran. P values and standard errors of mean were calculated by the qpAdm program applying a likelihood ratio test and the 5 cM block jackknifing method, respectively. No correction for multiple testing has been made. See also Extended Data Fig. 1 and Supplementary Tables 4–7. We further evaluated genetic heterogeneity with cladality tests using qpWave (Extended Data Fig. 1). Our results confirmed that various pairs within EBA Euboea, Aegina and Koufonisia were not cladal to each other with respect to a set of reference populations (model P < 0.05), highlighting substantial genetic variation among coeval groups. In stark contrast, in Early Middle Bronze Age (EMBA) Crete the rate of non-cladal pairs (25 of 741) was the one expected for true models of cladal pairs to be rejected with a cutoff of 5% given a uniform distribution of the P values. To increase the resolution of admixture inferences, we repeated qpAdm in groups of individuals ‘Crete Aposelemis N’ (n = 6), ‘S. Mainland-Islands LN-EBA’ (n = 13) and ‘Crete EMBA’ (n = 29) following a ‘competing’ approach described in previous studies (Methods and Supplementary Information 2). For Aposelemis, the one-way model from Neolithic western Anatolia was adequate when aceramic farmers from central Anatolia (Boncuklu site) were included in the reference populations but the one-way model with the latter as a source failed even without adding western Neolithic Anatolia to the references (P = 9.32 × 10−5) (Supplementary Note 2). Subsequently, we modelled the differences of the two descending ‘S. Mainland LN-EBA’ and ‘Crete EMBA’ groups from the earlier Aegean farmers with two-way models from these local farmers and various southwest Asian populations (Supplementary Table 7). Most of the two-way models including Neolithic Aposelemis were not rejected, indicating a decreased resolution owing to the low SNP coverage and small group size of Aposelemis. On the contrary, when models included ‘W. Anatolia N’ as a local source instead, only the one with an additional 28% contribution from the Eneolithic/BA Southern Caucasus was feasible for ‘S. Mainland-Islands LN-EBA’ (Fig. 3b). Accordingly, for ‘Crete EMBA’, the additional ancestry was better modelled with Late Chalcolithic/Early Bronze Age (LC-EBA) Anatolia (highest P = 4.9 × 10−3); however this model only became adequate as a three-way with an additional minute component (5%) from ‘W. Iran N’ (Fig. 3b). |
|

