Post by Admin on Nov 19, 2019 4:38:24 GMT
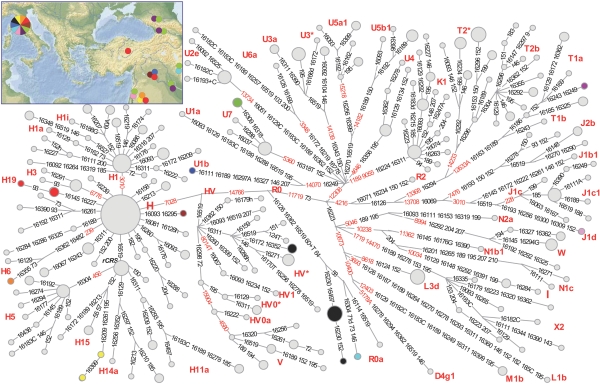
An Etruscan Origin in Anatolia?
Going back to the issue of the Etruscans’ origins, if the genetic resemblance between Turks and Tuscans reflects a common origin just before the onset of the Etruscan culture, as hypothesized by Herodotus and as considered in some recent studies [2], [6], [18], we would expect that the two populations separated about 3,000 years ago. To discriminate between the potentially similar effects of remote common origin and recent gene flow, we ran four independent analyses based on the IM method [19], [20]. In the model we tested, the two populations originate from a common ancestor, and may or may not exchange migrants after the split (Figure S7A). Assuming an average generation time of 25 years [16], [21] and no migration after the split from the common ancestors, the most likely separation time between Tuscany and Western Anatolia falls around 7,600 years ago, with a 95% credible interval between 5,000 and 10,000 (Figure 5). These results are robust to changes in the proportion of members of the initial population being ancestral to the two modern populations (Figure S7B). We also considered an expanded Anatolian sample (total sample size = 123 [11], [22]) coming from all over Turkey, to test whether a founder effect might have enhanced the role of the genetic drift in the previous analysis, inflating the divergence time estimates; the resulting distributions of separation times completely overlapped with those previously estimated, with a lower bound of the 95% credible interval never smaller than 5,300 years ago (Figure 5).

Figure 5. Separation time estimated by the IM model.
Estimation of the separation time between the gene pools of Anatolians (whether only Western Anatolians, or the expanded sample) and contemporary Tuscans (Casentino and Volterra). Means, upper bound and lower bound of the 95% credible intervals in four independent runs, obtained fixing the migration rate (indicated by dashed arrows) at 0, with mutation rate = 0.003 and assuming that the proportion of the ancestral population is equal in each descendant population (i.e. s = 0.5). Each analysis consisted of five coupled Markov chains, and 10,000,000 steps. Any degree of gene flow between the ancestors of Anatolians and Tuscans results in an increase of the estimate of the time since the population separation.
For these tests we chose the mutation rate (μ) estimated from the data in the previous ABC analyses (very close to the figure accounting for the time-dependency of the mitochondrial molecular clock [13], μ = 0.003). Tests were also run using the value incorporating a correction for the effects of purifying selection [23] (μ = 0.0014), always finding that it results in a further increase of the estimated separation times (Figure S7B). Only assuming very high mutation rates, at least twice as large as estimated in Henn et al. [13], was it possible to obtain separation times <5,000 years (Figure S7B). With both Anatolian samples, any degree of gene flow after separation between the ancestors of Tuscans and Anatolians resulted in more remote separation times.

Discussion
MtDNA data give much stronger support to a model of genetic continuity between the Etruscans and some Tuscans than to any other model tested, characterized by plausible population sizes and mutation rates. However, this clear picture emerges only when modern Tuscan communities are separately considered, highlighting the importance of population structure even at the small geographical scale. In a previous analysis of smaller samples we found no evidence of genealogical continuity since Etruscan times [5]. In this study, the larger sample sizes allowed us to separately investigate the relationships of each modern population with the Etruscans. A model of genealogical continuity across 2,500 years thus proved to best fit the observed data for Volterra, and especially Casentino, but not for another community dwelling in an area also rich with Etruscan archaeological remains (Murlo), nor (as expected) for the bulk of the current Tuscan population, here represented by a forensic sample of the inhabitants of Florence. Therefore, the present analysis indicates that the Etruscan genetic heritage is still present, but only in some isolates, whereas current Tuscans are not generally descended from Etruscan ancestors along the female lines. It also shows that there is no necessary correlation between the presence of archaeological remains and the biological roots of the inhabitants of the areas where these remains occur. Because Medieval Tuscans appears directly descended from Etruscan ancestors, one can reasonably speculate that the genetic build-up of the Murlo and Florence populations was modified by immigration in the last five centuries.
As for the second question, the IM analysis shows that indeed there might have been a genealogical link between modern Tuscans and the inhabitants of what Herodotus considered the Etruscans’ homeland, Western Anatolia. However, even under the unrealistic assumption of complete reciprocal isolation for millennia, the likely separation of the Tuscan and Anatolian gene pools must be placed long before the onset of the Etruscan culture, at least in Neolithic times; if isolation was incomplete, the estimated separation must be placed further back in time. Consistent with this view is the observation that Etruscan and Neolithic mtDNAs are close to each other in the two-dimensional plot of Figure S4C; however, a formal test would be necessary to draw firm conclusions from the simple observation of a genetic similarity. Separation times were very close when estimated both using a sample from Western Anatolia, and an expanded sample including individuals from much of Anatolia, and so the choice of the Anatolian population does not seem to affect the results of this analysis.
A general problem in ancient human DNA studies is the quality of the data; errors resulting from contamination, or from poor preservation of DNA in the specimens, are common. However, there are several reasons to be confident that the Etruscan sequences obtained in this study are authentic: (i) bones were recovered from burials according to the most stringent existing procedures and sent directly to the ancient DNA laboratory without manipulations; (ii) the mtDNA HVR-I motifs of the people who came in contact with the bones at any stage of the analysis do not match those obtained from the ancient samples (Table S1); (iii) the ancient samples were typed following the most stringent standard criteria for ancient DNA authentication; (iv) we used two different sequence determination procedures (classical methodology and high throughput methodology) and the results obtained from different extractions and different sequencing methodologies are concordant except in the regions of homopolymeric strings ≥5 bp that are problematic for the 454 pyrosequencing technology; in these cases, consensus sequences were determined considering only the results of the standard sequencing procedure; (v) sequences make phylogenetic sense, i.e. do not appear to be combinations of different sequences, possibly suggesting contamination by exogenous DNA.
Using such ancient DNA data for testing complex evolutionary models has become possible with the development of ABC and other recent Bayesian inference methods [24], [25]. These models, albeit more articulate than those that can be tested otherwise, are still a necessarily schematic representation of the processes affecting populations in the course of millennia. Many phenomena that could not be incorporated in the models, such as immigration from other sources or additional demographic fluctuations, most likely occurred and left a mark in the patterns of genetic diversity. In addition, specific phenomena may have involved mostly or exclusively males, resulting in genetic changes that are not recorded in mtDNA variation. Still, if we rule out the unlikely hypothesis that the Etruscans’ and their descendants’ population history was radically different for males and females, the picture emerging from this study is rather clear. The additional tests we ran (Type I error, Table 3) show that, at these sample sizes, we had a high probability to identify the correct evolutionary model.
As also suggested by the analysis of skull diversity [26], contacts between people from the Eastern Mediterranean shores and Central Italy likely date back to a remote stage of prehistory, possibly to the spread of farmers from the Near East during the Neolithic period [27], [28], but not necessarily so (we only estimated a minimum separation time between gene pools). At any rate, these contacts occurred much earlier than, and hence appear unrelated with, the onset of the Etruscan culture (Figure 5). We conclude that no available genetic evidence suggests an Etruscan origin outside Italy. While their culture disappeared from the records, the Etruscans’ mtDNAs did not; traces of this heritage are still recognizable. However, most current inhabitants of the ancient Etruscan homeland appear descended from different ancestors along the female lines, as clearly shown by the analysis of the urban (Florence) sample. Genetic continuity since the Etruscan’s time is still evident only in relatively isolated localities, such as Casentino and Volterra.
Ghirotto S, Tassi F, Fumagalli E, Colonna V, Sandionigi A, Lari M, et al. (2013) Origins and Evolution of the Etruscans’ mtDNA. PLoS ONE 8(2): e55519.
Going back to the issue of the Etruscans’ origins, if the genetic resemblance between Turks and Tuscans reflects a common origin just before the onset of the Etruscan culture, as hypothesized by Herodotus and as considered in some recent studies [2], [6], [18], we would expect that the two populations separated about 3,000 years ago. To discriminate between the potentially similar effects of remote common origin and recent gene flow, we ran four independent analyses based on the IM method [19], [20]. In the model we tested, the two populations originate from a common ancestor, and may or may not exchange migrants after the split (Figure S7A). Assuming an average generation time of 25 years [16], [21] and no migration after the split from the common ancestors, the most likely separation time between Tuscany and Western Anatolia falls around 7,600 years ago, with a 95% credible interval between 5,000 and 10,000 (Figure 5). These results are robust to changes in the proportion of members of the initial population being ancestral to the two modern populations (Figure S7B). We also considered an expanded Anatolian sample (total sample size = 123 [11], [22]) coming from all over Turkey, to test whether a founder effect might have enhanced the role of the genetic drift in the previous analysis, inflating the divergence time estimates; the resulting distributions of separation times completely overlapped with those previously estimated, with a lower bound of the 95% credible interval never smaller than 5,300 years ago (Figure 5).
Figure 5. Separation time estimated by the IM model.
Estimation of the separation time between the gene pools of Anatolians (whether only Western Anatolians, or the expanded sample) and contemporary Tuscans (Casentino and Volterra). Means, upper bound and lower bound of the 95% credible intervals in four independent runs, obtained fixing the migration rate (indicated by dashed arrows) at 0, with mutation rate = 0.003 and assuming that the proportion of the ancestral population is equal in each descendant population (i.e. s = 0.5). Each analysis consisted of five coupled Markov chains, and 10,000,000 steps. Any degree of gene flow between the ancestors of Anatolians and Tuscans results in an increase of the estimate of the time since the population separation.
For these tests we chose the mutation rate (μ) estimated from the data in the previous ABC analyses (very close to the figure accounting for the time-dependency of the mitochondrial molecular clock [13], μ = 0.003). Tests were also run using the value incorporating a correction for the effects of purifying selection [23] (μ = 0.0014), always finding that it results in a further increase of the estimated separation times (Figure S7B). Only assuming very high mutation rates, at least twice as large as estimated in Henn et al. [13], was it possible to obtain separation times <5,000 years (Figure S7B). With both Anatolian samples, any degree of gene flow after separation between the ancestors of Tuscans and Anatolians resulted in more remote separation times.
Discussion
MtDNA data give much stronger support to a model of genetic continuity between the Etruscans and some Tuscans than to any other model tested, characterized by plausible population sizes and mutation rates. However, this clear picture emerges only when modern Tuscan communities are separately considered, highlighting the importance of population structure even at the small geographical scale. In a previous analysis of smaller samples we found no evidence of genealogical continuity since Etruscan times [5]. In this study, the larger sample sizes allowed us to separately investigate the relationships of each modern population with the Etruscans. A model of genealogical continuity across 2,500 years thus proved to best fit the observed data for Volterra, and especially Casentino, but not for another community dwelling in an area also rich with Etruscan archaeological remains (Murlo), nor (as expected) for the bulk of the current Tuscan population, here represented by a forensic sample of the inhabitants of Florence. Therefore, the present analysis indicates that the Etruscan genetic heritage is still present, but only in some isolates, whereas current Tuscans are not generally descended from Etruscan ancestors along the female lines. It also shows that there is no necessary correlation between the presence of archaeological remains and the biological roots of the inhabitants of the areas where these remains occur. Because Medieval Tuscans appears directly descended from Etruscan ancestors, one can reasonably speculate that the genetic build-up of the Murlo and Florence populations was modified by immigration in the last five centuries.
As for the second question, the IM analysis shows that indeed there might have been a genealogical link between modern Tuscans and the inhabitants of what Herodotus considered the Etruscans’ homeland, Western Anatolia. However, even under the unrealistic assumption of complete reciprocal isolation for millennia, the likely separation of the Tuscan and Anatolian gene pools must be placed long before the onset of the Etruscan culture, at least in Neolithic times; if isolation was incomplete, the estimated separation must be placed further back in time. Consistent with this view is the observation that Etruscan and Neolithic mtDNAs are close to each other in the two-dimensional plot of Figure S4C; however, a formal test would be necessary to draw firm conclusions from the simple observation of a genetic similarity. Separation times were very close when estimated both using a sample from Western Anatolia, and an expanded sample including individuals from much of Anatolia, and so the choice of the Anatolian population does not seem to affect the results of this analysis.
A general problem in ancient human DNA studies is the quality of the data; errors resulting from contamination, or from poor preservation of DNA in the specimens, are common. However, there are several reasons to be confident that the Etruscan sequences obtained in this study are authentic: (i) bones were recovered from burials according to the most stringent existing procedures and sent directly to the ancient DNA laboratory without manipulations; (ii) the mtDNA HVR-I motifs of the people who came in contact with the bones at any stage of the analysis do not match those obtained from the ancient samples (Table S1); (iii) the ancient samples were typed following the most stringent standard criteria for ancient DNA authentication; (iv) we used two different sequence determination procedures (classical methodology and high throughput methodology) and the results obtained from different extractions and different sequencing methodologies are concordant except in the regions of homopolymeric strings ≥5 bp that are problematic for the 454 pyrosequencing technology; in these cases, consensus sequences were determined considering only the results of the standard sequencing procedure; (v) sequences make phylogenetic sense, i.e. do not appear to be combinations of different sequences, possibly suggesting contamination by exogenous DNA.
Using such ancient DNA data for testing complex evolutionary models has become possible with the development of ABC and other recent Bayesian inference methods [24], [25]. These models, albeit more articulate than those that can be tested otherwise, are still a necessarily schematic representation of the processes affecting populations in the course of millennia. Many phenomena that could not be incorporated in the models, such as immigration from other sources or additional demographic fluctuations, most likely occurred and left a mark in the patterns of genetic diversity. In addition, specific phenomena may have involved mostly or exclusively males, resulting in genetic changes that are not recorded in mtDNA variation. Still, if we rule out the unlikely hypothesis that the Etruscans’ and their descendants’ population history was radically different for males and females, the picture emerging from this study is rather clear. The additional tests we ran (Type I error, Table 3) show that, at these sample sizes, we had a high probability to identify the correct evolutionary model.
As also suggested by the analysis of skull diversity [26], contacts between people from the Eastern Mediterranean shores and Central Italy likely date back to a remote stage of prehistory, possibly to the spread of farmers from the Near East during the Neolithic period [27], [28], but not necessarily so (we only estimated a minimum separation time between gene pools). At any rate, these contacts occurred much earlier than, and hence appear unrelated with, the onset of the Etruscan culture (Figure 5). We conclude that no available genetic evidence suggests an Etruscan origin outside Italy. While their culture disappeared from the records, the Etruscans’ mtDNAs did not; traces of this heritage are still recognizable. However, most current inhabitants of the ancient Etruscan homeland appear descended from different ancestors along the female lines, as clearly shown by the analysis of the urban (Florence) sample. Genetic continuity since the Etruscan’s time is still evident only in relatively isolated localities, such as Casentino and Volterra.
Ghirotto S, Tassi F, Fumagalli E, Colonna V, Sandionigi A, Lari M, et al. (2013) Origins and Evolution of the Etruscans’ mtDNA. PLoS ONE 8(2): e55519.