|
|
Post by Admin on Feb 24, 2020 23:35:52 GMT
Sampling DNA from ancient remains allows scientists to get a snapshot of people living at a specific time and place, instead of using modern DNA and inferring the past based on assumptions and mathematical models. When the team compared the DNA of 70 ancient individuals collected from Sardinia to the DNA of other ancient and modern individuals, they uncovered two major patterns. First, they saw that Sardinian individuals in the Middle Neolithic period (4100-3500 BCE) were closely related to people from mainland Europe of the time. Genetic ancestry then remained relatively stable on the island through at least the end of the "Nuragic" period (~900 BCE). This pattern differs from other regions of mainland Europe which experienced new ancestries entering from people moving across the continent in the Bronze Age.  The results also show the development of Sardinia's distinctive nuraghe stone towers and culture (after which the Nuragic period is named) did not coincide with any detectable, new genetic ancestry arriving to the island. "We found striking stability in ancestry from the Middle Neolithic through the end of the Nuragic period in Sardinia," said Joe Marcus, a Ph.D. student in the Department of Human Genetics at UChicago and a co-first author on the paper. Second, the team found evidence of the arrival of different populations across the Mediterranean, first with Phoenicians originating from the Levant (modern-day Lebanon) and Punics, whose culture centered in Carthage (modern-day Tunisia). Then, new ancestry continued to appear during the Roman period and further in the Medieval period, as Sardinia became historically influenced by migration of people from modern-day Italy and Spain. "We observed clear signals of dynamic periods of contact linking the island to the rest of the Mediterranean, appearing first in individuals from two Phoenician and Punic sites as early as 500 BCE, and then in individuals from the Roman and Medieval periods," said Harald Ringbauer, Ph.D., a postdoctoral researcher involved in the computational data analysis at UChicago and a co-first author on the paper. The group's results help explain similarities with DNA from mainland European individuals of the Neolithic and Copper Age, such as "Ötzi the Iceman," an almost perfectly preserved, 5,300-year-old human discovered in alpine ice in northern Italy in 1991. Specifically, among modern Europeans, Ötzi's DNA is most similar to modern-day Sardinians. The new study supports the theory that this similarity remains because Sardinia had less turnover of genetic ancestry over time than mainland Europe, which experienced large-scale migrations in the Bronze Age.  Insights from the past, implications for the present Besides providing new insight into mysteries of the past, studying ancient DNA also has implications for the well-being of present-day humans. This model of Sardinia's population history—establishment followed by relative isolation and then the arrival of new sources of diversity—provides a new framework for understanding how genetic variants with health implications became more frequent on the island. "For future studies, we want to look more precisely at mutations that we think are involved in disease to see in which period they changed in frequency and how quickly they changed," Novembre said. "That will help us understand the processes acting on these diseases, and in turn gain a richer view that may yield insights for human health." The study, "Genetic history from the Middle Neolithic to present on the Mediterranean island of Sardinia," was published February 24, 2020 in Nature Communications. An independent study in Nature Ecology and Evolution also published February 24, 2020 comes to similar conclusions using different samples. |
|
|
|
Post by Admin on Feb 25, 2020 21:01:14 GMT
Abstract The island of Sardinia has been of particular interest to geneticists for decades. The current model for Sardinia’s genetic history describes the island as harboring a founder population that was established largely from the Neolithic peoples of southern Europe and remained isolated from later Bronze Age expansions on the mainland. To evaluate this model, we generate genome-wide ancient DNA data for 70 individuals from 21 Sardinian archaeological sites spanning the Middle Neolithic through the Medieval period. The earliest individuals show a strong affinity to western Mediterranean Neolithic populations, followed by an extended period of genetic continuity on the island through the Nuragic period (second millennium BCE). Beginning with individuals from Phoenician/Punic sites (first millennium BCE), we observe spatially-varying signals of admixture with sources principally from the eastern and northern Mediterranean. Overall, our analysis sheds light on the genetic history of Sardinia, revealing how relationships to mainland populations shifted over time.  Introduction The whole-genome sequencing in 2012 of “Ötzi”, an individual who was preserved in ice for over 5000 years near the Italo-Austrian border, revealed a surprisingly high level of shared ancestry with present-day Sardinian individuals1,2. Subsequent work on genome-wide variation in ancient Europeans found that most “early European farmer” individuals, even when from geographically distant locales (e.g., from Sweden, Hungary and Spain) have their highest genetic affinity with present-day Sardinian individuals3,4,5,6. Accumulating ancient DNA (aDNA) results have provided a framework for understanding how early European farmers show such genetic affinity to modern Sardinians. In this framework, Europe was first inhabited by Paleolithic and later Mesolithic hunter-gatherer groups. Then, starting about 7000 BCE, farming peoples arrived from the Middle East as part of a Neolithic transition7, spreading through Anatolia and the Balkans8,9 while progressively admixing with local hunter-gatherers10. Major movements from the Eurasian Steppe, beginning about 3000 BCE, resulted in further admixture throughout Europe11,12,13,14. These events are typically modeled in terms of three ancestry components: western hunter gatherers (“WHG”), early European farmers (“EEF”), and Steppe pastoralists (“Steppe”). Within this broad framework, the island of Sardinia is thought to have received a high level of EEF ancestry early on and then remained mostly isolated from the subsequent admixture occurring on mainland Europe1,2. However, this specific model for Sardinian population history has not been tested with genome-wide aDNA data from the island. The oldest known human remains on Sardinia date to ~20,000 years ago15. Archeological evidence suggests Sardinia was not densely populated in the Mesolithic, and experienced a population expansion coinciding with the Neolithic transition in the sixth millennium BCE16. Around this time, early Neolithic pottery assemblages were spreading throughout the western Mediterranean, including Sardinia, in particular vessels decorated with Cardium shell impressions (variably described as Impressed Ware, Cardial Ware, Cardial Impressed Ware)17, with radiocarbon dates indicating a rapid westward maritime expansion around 5500 BCE18. In the later Neolithic, obsidian originating from Sardinia is found throughout many western Mediterranean archeological sites19, indicating that the island was integrated into a maritime trade network. In the middle Bronze Age, about 1600 BCE, the “Nuragic” culture emerged, named for the thousands of distinctive stone towers, known as nuraghi20. During the late Nuragic period, the archeological and historical record shows the direct influence of several major Mediterranean groups, in particular the presence of Mycenaean, Levantine and Cypriot traders. The Nuragic settlements declined throughout much of the island as, in the late 9th and early 8th century BCE, Phoenicians originating from present-day Lebanon and northern Palestine established settlements concentrated along the southern shores of Sardinia21. In the second half of the 6th century BCE, the island was occupied by Carthaginians (also known as Punics), expanding from the city of Carthage on the North-African coast of present-day Tunisia, which was founded in the late 9th century by Phoenicians22,23. Sardinia was occupied by Roman forces in 237 BCE, and turned into a Roman province a decade later24. Throughout the Roman Imperial period, the island remained closely aligned with both Italy and central North Africa. After the fall of the Roman empire, Sardinia became increasingly autonomous24, but interaction with the Byzantine Empire, the maritime republics of Genova and Pisa, the Catalan and Aragonese Kingdom, and the Duchy of Savoy and Piemonte continued to influence the island25,26. The population genetics of Sardinia has long been studied, in part because of its importance for medical genetics27,28. Pioneering studies found evidence that Sardinia is a genetic isolate with appreciable population substructure29,30,31. Recently, Chiang et al.32 analyzed whole-genome sequences33 together with continental European aDNA. Consistent with previous studies, they found the mountainous Ogliastra region of central/eastern Sardinia carries a signature of relative isolation and subtly elevated levels of WHG and EEF ancestry. Four previous studies have analyzed aDNA from Sardinia using mitochondrial DNA. Ghirotto et al.34 found evidence for more genetic turnover in Gallura (a region in northern Sardinia with cultural/linguistic connections to Corsica) than Ogliastra. Modi et al.35 sequenced mitogenomes of two Mesolithic individuals and found support for a model of population replacement in the Neolithic. Olivieri et al.36 analyzed 21 ancient mitogenomes from Sardinia and estimated the coalescent times of Sardinian-specific mtDNA haplogroups, finding support for most of them originating in the Neolithic or later, but with a few coalescing earlier. Finally, Matisoo-Smith et al.37 analyzed mitogenomes in a Phoenician settlement on Sardinia and inferred continuity and exchange between the Phoenician population and broader Sardinia. One additional study recovered β-thalessemia variants in three aDNA samples and found one carrier of the cod39 mutation in a necropolis used in the Punic and Roman periods38. Despite the initial insights these studies reveal, none of them analyze genome-wide autosomal data, which has proven to be useful for inferring population history39.  Here, we generate genome-wide data from the skeletal remains of 70 Sardinian individuals radiocarbon dated to between 4100 BCE and 1500 CE. We investigate three aspects of Sardinian population history: First, the ancestry of individuals from the Sardinian Neolithic (ca. 5700–3400 BCE)—who were the early peoples expanding onto the island at this time? Second, the genetic structure through the Sardinian Chalcolithic (i.e., Copper Age, ca. 3400–2300 BCE) to the Sardinian Bronze Age (ca. 2300–1000 BCE)—were there genetic turnover events through the different cultural transitions observed in the archeological record? And third, the post-Bronze Age contacts with major Mediterranean civilizations and more recent Italian populations—have they resulted in detectable gene flow? Our results reveal insights about each of these three periods of Sardinian history. Specifically, our earliest samples show affinity to the early European farmer populations of the mainland, then we observe a period of relative isolation with no significant evidence of admixture through the Nuragic period, after which we observe evidence for admixture with sources from the northern and eastern Mediterranean. |
|
|
|
Post by Admin on Feb 26, 2020 19:59:11 GMT
Results Ancient DNA from Sardinia We organized a collection of skeletal remains (Supp. Fig. 1) from (1) a broad set of previously excavated samples initially used for isotopic analysis40, (2) the Late Neolithic to Bronze Age Seulo cave sites of central Sardinia41, (3) the Neolithic Sites Noedalle and S’isterridolzu42, (4) the Phoenician-Punic sites of Monte Sirai23 and Villamar43, (5) the Imperial Roman period site at Monte Carru (Alghero)44, (6) medieval remains from the site of Corona Moltana45, (7) medieval remains from the necropolis of the Duomo of San Nicola46. We sequenced DNA libraries enriched for the complete mitochondrial genome as well as a targeted set of 1.2 million single nucleotide polymorphisms (SNPs)47. After quality control, we arrived at a final set of 70 individuals with an average coverage of 1.02× at targeted SNPs (ranging from 0.04× to 5.39× per individual) and a median number of 466,049 targeted SNPs covered at least once per individual. We obtained age estimates by either direct radiocarbon dating (n = 53), previously reported radiocarbon dates (n = 13), or archeological context and radiocarbon dates from the same burial site (n = 4). The estimated ages range from 4100 years BCE to 1500 years CE (Fig. 1, Supp. Data 1A). We pragmatically grouped the data into broad periods: Middle/Late Neolithic (‘Sar-MN’, 4100–3500 BCE, n = 6), Early Copper Age (‘Sar-ECA’, 3500–2500 BCE, n = 3), Early Middle Bronze Age (‘Sar-EMBA’, 2500–1500 BCE, n = 27), and Nuragic (‘Sar-Nur’, 1500–900 BCE, n = 16). For the post-Nuragic sites, there is substantial genetic heterogeneity within and among sites, and so we perform analysis per site when grouping is necessary (‘Sar-MSR’ and ‘Sar-VIL’ for the Phoenician and Punic sites of Monte Sirai, n = 2; and Villamar, n = 6; ‘Sar-ORC002’ for a Punic period individual from the interior site of S’Orcu ’e Tueri, n = 1; ‘Sar-AMC’ for the Roman period site of Monte Carru near Alghero, n = 3; ‘Sar-COR’ for the early medieval individuals from the site of Corona Moltana, n = 2; and ‘Sar-SNN’ for the medieval San Nicola necropoli, n = 4). Figure 1 provides an overview of the sample. 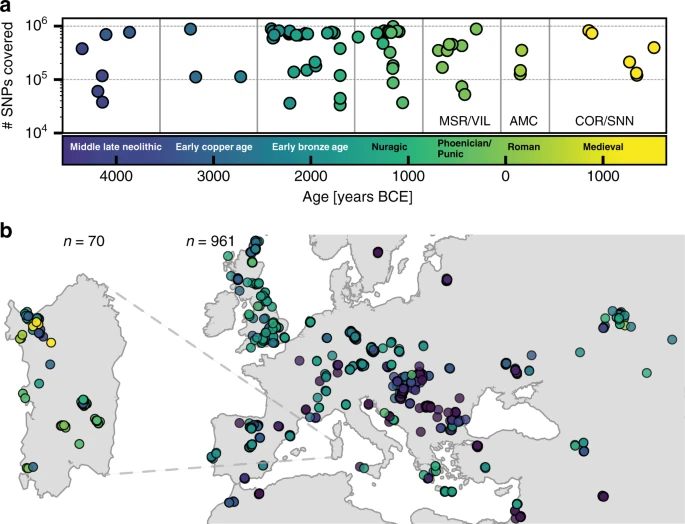 Fig. 1: Number of SNPs covered, sampling locations and ages of ancient individuals. To assess the relationship of the ancient Sardinian individuals to other ancient and present-day west Eurasian and North-African populations we analyzed our individuals alongside published autosomal DNA data (ancient: 972 individuals9,10,13,48,49,50; modern: 1963 individuals from outside Sardinia7 and 1577 individuals from Sardinia32,33). For some analyses, we grouped the modern Sardinian individuals into eight geographic regions (see inset in panel c of Fig. 2 for listing and abbreviations, also see Supp. Data 1E) and for others we subset the more isolated Sardinian region of Ogliastra (‘Sar-Ogl’, n = 419) and the remainder (‘Sar-non Ogl’, n = 1158). As with other human genetic variation studies, population annotations are important to consider in the interpretation of results. 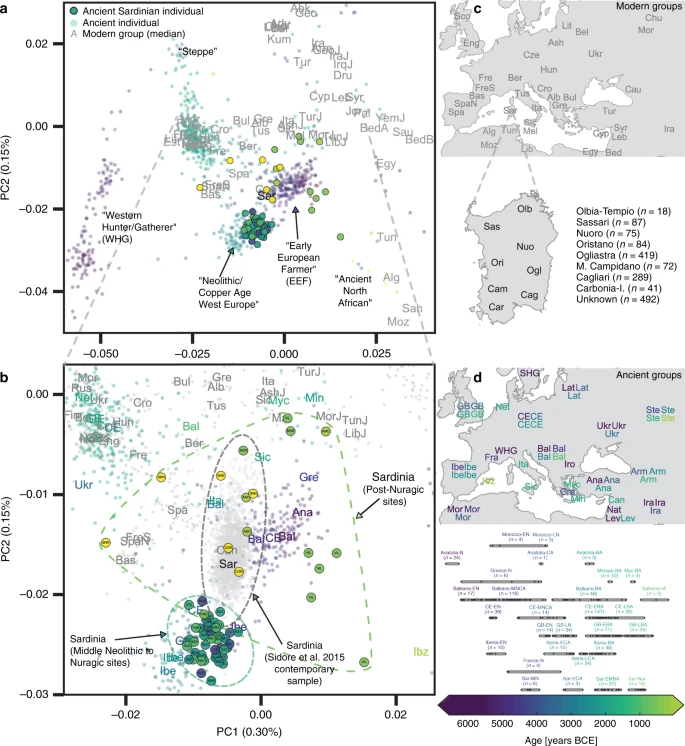 Fig. 2: Principal components analysis based on the Human Origins dataset. a Projection of ancient individuals’ genotypes onto principal component axes defined by modern Western Eurasians and North Africans (gray labels, see panel (c) for legend for all abbreviations but ‘Can', for Canary Islands). b Zoom into the region most relevant for Sardinia. Each projected ancient individual is displayed as a transparent colored point in panel (a) and (b), with the color determined by the age of each sample (see panel (d) for legend). In panel (b), median PC1 and PC2 values for each population are represented by three-letter abbreviations, with black or gray font for moderns and a color-coded font based on the mean age for ancient populations. Ancient Sardinian individuals are plotted as circles with edges, color-coded by age, and with the first three letters of their sample ID (which typically indicates the archeological site). Modern individuals from the Sidore et al. sample of Sardinia are represented with gray circles and modern individuals from the reference panel with gray squares. See Fig. 5 for a zoomed in representation with detailed province labels for Sardinian individuals. The full set of labels and abbreviations are described in Supp. Data 1E, F. c Geographic legend of present-day individuals from the Human Origins and our Sardinian reference dataset. d Timeline of selected ancient groups. Note: The same geographic abbreviation can appear multiple times with different colors to represent groups with different median ages. |
|
|
|
Post by Admin on Feb 27, 2020 0:25:30 GMT
Similarity to western mainland Neolithic populations We found low differentiation between Middle/Late Neolithic Sardinian individuals and Neolithic western mainland European populations, in particular groups from Spain (Iberia-EN) and southern France (France-N). When projecting ancient individuals onto the top two principal components (PCs) defined by modern variation, the Neolithic ancient Sardinian individuals sit between early Neolithic Iberian and later Copper Age Iberian populations, roughly on an axis that differentiates WHG and EEF populations, and embedded in a cluster that additionally includes Neolithic British individuals (Fig. 2). This result is also evident in terms of genetic differentiation, with low pairwise FST ≈ 0.005–0.008, between Middle/Late Neolithic and Neolithic western mainland European populations (Fig. 3). Pairwise outgroup-f3 analysis shows a similar pattern, with the highest values of f3 (i.e., most shared drift) being with Western European Neolithic and Copper Age populations (Fig. 3), gradually dropping off for populations more distant in time or space (Supp. Fig. 10). 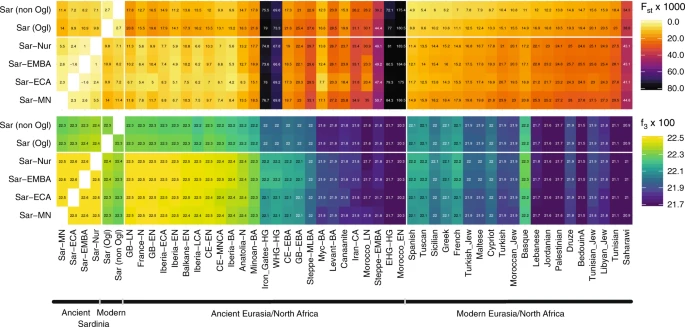 Fig. 3: Genetic similarity matrices. Ancient Sardinian individuals are shifted towards WHG individuals in the top two PCs relative to early Neolithic Anatolians (Fig. 2). Analysis using qpAdm shows that a two-way admixture model between WHG and Neolithic Anatolian populations is consistent with our data (e.g., p = 0.376 for Sar-MN, Table 1), similar to other western European populations of the early Neolithic (Supp. Table 1). The method estimates ancient Sardinian individuals harbor HG ancestry (≈17 ± 2%) that is higher than early Neolithic mainland populations (including Iberia, 8.7 ± 1.1%), but lower than Copper Age Iberians (25.1 ± 0.9%) and about the same as Southern French Middle-Neolithic individuals (21.3 ± 1.5%, Table 1, Supp. Fig. 13, ± denotes plus and minus one standard error). In explicit models of continuity (using qpAdm, see Methods) the southern French Neolithic individuals (France-N) are consistent with being a single source for Middle/Late Neolithic Sardinia (p = 0.38 to reject the model of one population being the direct source of the other); followed by other western populations high in EEF ancestry, though with poor fit (qpAdm p-values < 10−5, Supp. Table 2). France-N may result in improved fits as it is a better match for the WHG and EEF proportions seen in Middle/Late Neolithic Sardinia (Supp. Table 1). As we discuss below, caution is necessary as there is a lack of aDNA from other relevant populations of the same period (such as mainland Italian Neolithic cultures and neighboring islands). For our sample from the Middle Neolithic through the Nuragic (n = 52 individuals), we were able to infer mtDNA haplotypes for each individual and Y haplotypes for 30 out of 34 males. The mtDNA haplotypes belong to macro-haplogroups HV (n = 20), JT (n = 19), U (n = 12), and X (n = 1), a composition broadly similar to other European Neolithic populations. For Y haplotypes, we found at least one carrier for each of three major Sardinia-specific Y founder clades (within the haplogroups I2-M26, G2-L91, and R1b-V88) that were identified previously based on modern Sardinian data51. More than half of the 31 identified Y haplogroups were R1b-V88 or I2-M223 (n = 11 and 8, respectively, Supp. Fig. 6, Supp. Data 1B), both of which are also prevalent in Neolithic Iberians14. Compared with most other ancient populations in our reference dataset, the frequency of R1b-V88 (Supp. Note 3, Supp. Fig. 6) is relatively high, but as we observed clustering of Y haplogroups by sample location (Supp. Data 1B) caution should be exercised with interpreting our results as estimates for island-wide Y haplogroup frequencies. The oldest individuals in our reference data carrying R1b-V88 or I2-M223 were Balkan hunter-gatherer and Neolithic individuals, and both haplogroups later appear also in western Neolithic populations (Supp. Figs. 7–9). |
|
|
|
Post by Admin on Feb 27, 2020 22:15:04 GMT
From the Nuragic period to present-day Sardinia: signatures of admixture We found multiple lines of evidence for gene flow into Sardinia after the Nuragic period. The present-day Sardinian individuals from the Sidore et al. sample are shifted from the Nuragic period ancients on the western Eurasian/North-African PCA (Fig. 2). Using a “shrinkage” correction method for the projection is key for detecting this shift (see Supp. Fig. 23 for an evaluation of different PCA projection techniques). In the ADMIXTURE results (Fig. 4), present-day Sardinian individuals carry a modest “Steppe-like” ancestry component (but generally less than continental present-day European populations), and an appreciable “eastern Mediterranean” ancestry component (also inferred at a high fraction in other present-day Mediterranean populations, such as Sicily and Greece) relative to Nuragic period and earlier Sardinian individuals. 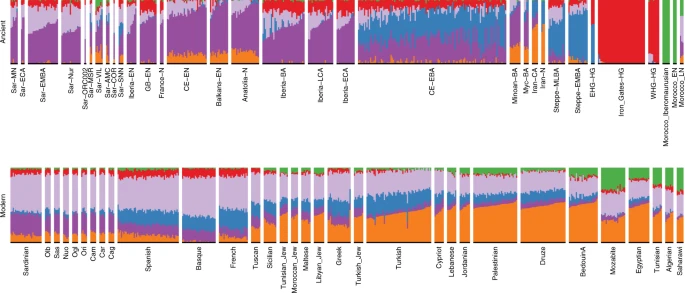 Fig. 4: Admixture coefficients estimated by ADMIXTURE (K = 6). To further refine this recent admixture signal, we considered two-way, three-way, and four-way models of admixture with qpAdm (Table 2, Supp. Figs. 15–18, Supp. Tables 3–5). We find three-way models fit well (p > 0.01) that contain admixture between Nuragic Sardinia, one northern Mediterranean source (e.g., individuals with group labels Lombardy, Tuscan, French, Basque, Spanish) and one eastern Mediterranean source (e.g., individuals with group labels Turkish-Jew, Libyan-Jew, Maltese, Tunisian-Jew, Moroccan-Jew, Lebanese, Druze, Cypriot, Jordanian, Palestinian) (Table 2C, D). Maltese and Sicilian individuals can provide two-way model fits (Table 2B), but appear to reflect a mixture of N. Mediterranean and E. Mediterranean ancestries, and as such they can serve as single-source proxies in two-way admixture models with Nuragic Sardinia. For four-way models including N. African ancestry, the inferences of N. African ancestry are negligible (though as we show below, forms of N. African ancestry were already likely present in the eastern Mediterranean components). Because of limited sample sizes and ancestral source mis-specification, caution is warranted when interpreting inferred admixture fractions; however, the results indicate that complex post-Nuragic gene flow has likely played a role in the population genetic history of Sardinia. Refined signatures of post-Nuragic admixture and heterogeneity To more directly evaluate the models of post-Nuragic admixture, we obtained aDNA from 17 individuals sampled from post-Nuragic sites. The post-Nuragic individuals spread across a wide range of the PCA, and many shift towards the “eastern” and “northern” Mediterranean sources posited above (Fig. 2). We confidently reject qpAdm models of continuity from the Nuragic period for all of these post-Nuragic samples, apart from a sample from S’Orcu ’e Tueri (ORC002, Table 2E, Supp. Table 6). The ADMIXTURE results concur, most post-Nuragic individuals show the presence of novel ancestry components not inferred in any of the more ancient individuals (Fig. 4). Consistent with an influx of novel ancestry, we observed that haplogroup diversity increases after the Nuragic period. In particular, we identified one carrier of the mtDNA haplogroup L2a at both the Punic Villamar site and the Roman Monte Carru site. At present, this mtDNA haplogroup is common across Africa, but so far undetected in samples from Sardinia36. We also found several Y haplogroups absent in our Neolithic trough the Nuragic period sample (Supp. Fig. 6). R1b-M269, at about 15% within modern Sardinian males51, appears in one Punic (VIL011) and two Medieval individuals (SNN002 and SNN004). We also observed J1-L862 in one individual from a Punic site (VIL007) and E1b-L618 in one medieval individual (SNN001). Notably, J1-L862 first appears in Levantine Bronze Age individuals within the ancient reference dataset and is at about 5% frequency in Sardinia today. We used individual-level qpAdm models to further investigate the presence of these new ancestries (Supp. Data 3). In addition to the original Neolithic Anatolian (Anatolia-N) and Hunter Gatherer (WHG) sources that were sufficient to model ancient Sardinians through the Nuragic period, we fit models with representatives of Steppe (Steppe-EMBA), Neolithic Iranian (Iran-N), and Neolithic North-African (Morocco-EN) ancestry as sources. We observe the presence of the Steppe-EMBA (point estimates ranging 0–20%) and Iran-N components (point estimates ranging 0–25%) in many of the post-Nuragic individuals (Supp. Fig. 14). All six individuals from the Punic Villamar site were inferred to have substantial levels of ancient North-African ancestry (point estimates ranging 20–35%, Supp. Fig. 14, also see ADMIXTURE and PCA results, Figs. 2 and 4). When fit with the same five-way admixture model, present-day Sardinians have a small but detectable level of North-African ancestry (Supp. Fig. 14, also see ADMIXTURE analysis, Fig. 4). Models with direct continuity from Villamar to the present are rejected (Table 2F, Supp. Table 6). In contrast, nearly all the other post-Nuragic sites produce viable models as single sources for the modern Sardinians (e.g., Sar-COR qpAdm p-values of 0.16 and 0.261 for Cagliari and Ogliastra, respectively; Sar-SNN qpAdm p-values of 0.037 and 0.016, similarly Table 2F, Supp. Table 6). We found some evidence of substructure: Sar-ORC002 (from an interior site) is more consistent with being a single source for Ogliastra than Cagliari, whereas Sar-AMC shows an opposite pattern (Supp. Table 6). We also carried out three-way admixture models for each post-Nuragic Sardinian individual using the Nuragic sample as a source or outgroup, and potential sources from various ancient samples that are representative of different regions of the Mediterranean. We found a range of models can be fit for each individual (Supp. Tables 7–8). For the models with Nuragic as a source, by varying the proxy populations, one can obtain fitted models that vary widely in the inferred Nuragic component (e.g., individual COR002 has a range from 4.4 to 87.8% across various fitted models; similarly, individual AMC001, with North-African mtDNA haplogroup U6a, had a range form 0.2 to 43.1%, see Supp. Tables 7–8). The ORC002 sample had the strongest evidence of Nuragic ancestry (range from 62.8 to 96.3%, see Supp. Tables 7–8). Further, the VIL, MSR, and AMC individuals can be modeled with Nuragic Sardinian individuals included as a source or as an outgroup, while the two COR and ORC002 individuals can only be modeled with Nuragic individuals included as a source. One individual from the medieval period San Nicola Necropoli (SNN001) was distinct in that we found their ancestry can be modeled in a single-source model as descendant of a population represented by present-day Basque individuals (Supp. Table 8). When we apply the same approach to present-day Sardinian individuals, we find models with the Nuragic sample as an outgroup fail in most cases (Supp. Table 9). For models that include Nuragic as a possible source, each present-day individual is consistent with a wide range of Nuragic ancestry. The models with the largest p-values return fractions of Nuragic ancestry that are close to, or higher than 50% (Supp. Table 9), similar to observed in our population-level modeling (Table 2). |
|

