Source of ANE Ancestry in the LBA Khövsgöl Population.
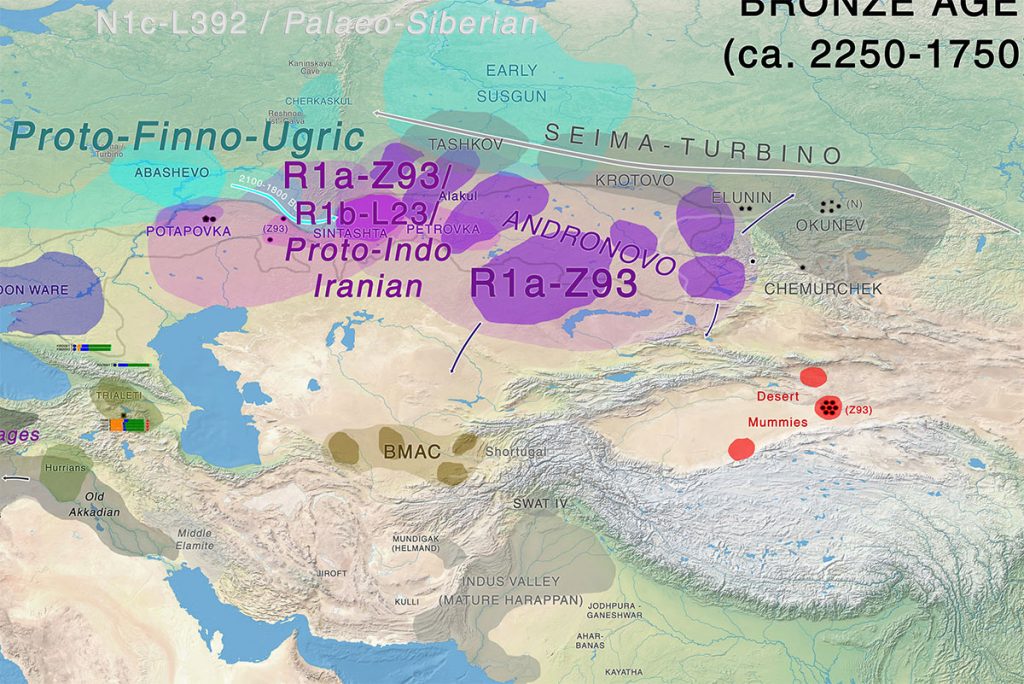
Previous studies show a close genetic relationship between WSH populations and ANE ancestry, as Yamnaya and Afanasievo are modeled as a roughly equal mixture of early Holocene Iranian/Caucasus ancestry (IRC) and Mesolithic Eastern European hunter-gatherers, the latter of which derive a large fraction of their ancestry from ANE (20, 28). It is therefore important to pinpoint the source of ANE-related ancestry in the Khövsgöl gene pool: that is, whether it derives from a pre-Bronze Age ANE population (such as the one represented by AG3) or from a Bronze Age WSH population that has both ANE and IRC ancestry. To test these competing hypotheses, we systematically compared various admixture models of the main cluster using the qpAdm program (20). Ancient Baikal populations were chosen as a proxy based on both their spatiotemporal and genetic similarities with the Khövsgöl main cluster (Figs. 2 and 3). When the early Neolithic Shamanka_EN is used as a proxy, we find that Baikal+ANE provides a better fit to the main cluster than Baikal+WSH, although no two-way admixture model provides a sufficient fit (P ≥ 0.05) (SI Appendix, Table S5). Adding a WSH population as the third source results in a sufficient three-way mixture model of Baikal+ANE+WSH with a small WSH contribution to the main cluster (e.g., P = 0.180 for Shamanka_EN+AG3+Sintashta with 3.7 ± 2.0% contribution from Sintashta) (Fig. 4 and SI Appendix, Table S6).

Fig. 4.
Admixture modeling of Altai populations and the Khövsgöl main cluster using qpAdm. For the archaeological populations, (A) Shamanka_EBA and (B and C) Khövsgöl, each colored block represents the proportion of ancestry derived from a corresponding ancestry source in the legend. Error bars show 1 SE. (A) Shamanka_EBA is modeled as a mixture of Shamanka_EN and AG3. The Khövsgöl main cluster is modeled as (B) a two-way admixture of Shamanka_EBA+Sintashta and (C) a three-way admixture Shamanka_EN+AG3+Sintashta. Details of the admixture models are provided in SI Appendix, Tables S5 and S6.
Using the temporally intermediate EBA population Shamanka_EBA, we can narrow down the time for the introduction of WSH ancestry into the main cluster. Shamanka_EBA is modeled well as a two-way mixture of Shamanka_EN and ANE (P = 0.158 for Shamanka_EN+AG3) (Fig. 4) but not as a mixture of Shamanka_EN and WSH (P ≤ 2.91 × 10−4) (SI Appendix, Table S5), suggesting no detectable WSH contribution through the early Bronze Age. Similar results are obtained for other Late Neolithic and EBA populations from the Baikal region (SI Appendix, Table S5). In contrast, the Khövsgöl main cluster is modeled well by Shamanka_EBA+WSH but not by Shamanka_EBA+ANE (P ≥ 0.073 and P ≤ 0.038, respectively) (SI Appendix, Table S5). A three-way model of Shamanka_EBA+ANE+WSH confirms this by providing the ANE contribution around zero (SI Appendix, Table S6). The amount of WSH contribution remains small (e.g., 6.4 ± 1.0% from Sintashta) (Fig. 4 and SI Appendix, Table S5). Assuming that the early Neolithic populations of the Khövsgöl region resembled those of the nearby Baikal region, we conclude that the Khövsgöl main cluster obtained ∼11% of their ancestry from an ANE source during the Neolithic period and a much smaller contribution of WSH ancestry (4–7%) beginning in the early Bronze Age.
Admixture Testing of Genetic Outliers.
Using the same approach, we obtained reasonable admixture models for the two outliers, ARS017 and ARS026. The eastern outlier ARS017, a female, shows an extra affinity with early Neolithic individuals from the Russian far east (Devil’s Gate) (22) and in general with contemporary East Asians (e.g., Han Chinese) compared with the Khövsgöl main cluster (Fig. 3B and SI Appendix, Fig. S12). ARS017 is also similar to Shamanka_EN in showing no significant difference in qpAdm (SI Appendix, Fig. S12 and Table S7). Using contemporary East Asian proxies, ARS017 is modeled as a mixture of predominantly Ulchi and a minor component (6.1–9.4%) that fits most ancient western Eurasian groups (P = 0.064–0.863) (SI Appendix, Table S7). This minor Western component may result from ANE ancestry; however, given the minimal western Eurasian contribution, we do not have sufficient power to accurately characterize this individual’s western Eurasian ancestry.
The Western outlier ARS026, a male dating to the end of the radiocarbon series, has the highest outgroup-f3 with the main LBA Khövsgöl cluster, with extra affinity toward Middle Bronze Age (MBA) individuals from the Sintashta culture (Fig. 3B and SI Appendix, Fig. S13) (1). DNA recovered from this individual exhibited expected aDNA damage patterns (SI Appendix, Fig. S5) but was otherwise excellently preserved with >47% endogenous content and very low estimated contamination (1% mitochondrial; 0.01% nuclear). ARS026 is well modeled as a two-way mixture of Shamanka_EBA and Sintashta (P = 0.307; 48.6 ± 2.0% from Sintashta) (SI Appendix, Table S7). Similar to ARS026, contemporaneous LBA Karasuk individuals from the Altai (1400–900 BCE) (1, 29) also exhibit a strong extra genetic affinity with individuals associated with the earlier Sintashta and Andronovo cultures (SI Appendix, Fig. S14). Although two-way admixture models do not fit (P ≤ 0.045) (SI Appendix, Table S8), the Karasuk can be modeled as a three-way mixture of Shamanka_EBA/Khövsgöl and AG3 and Sintashta, suggesting an eastern Eurasian source with slightly higher ANE ancestry than those used in our modeling (P ≥ 0.186) (SI Appendix, Table S8). Like ARS026, admixture coefficients for the Karasuk suggest that MBA/LBA groups like the Sintashta or Srubnaya are a more likely source of their WSH ancestry than the EBA groups, like the Yamnaya or Afanasievo. Notably, Karasuk individuals are extremely heterogeneous in their genetic composition, with the genetically easternmost Eurasian individual nearly overlapping with the EBA Baikal groups (Fig. 2A and SI Appendix, Figs. S7 and S8). Earlier groups, such as the Afanasievo, Sintashta, and Andronovo, are mostly derived from WSH ancestries, and this may suggest that admixture in the Altai-Sayan region only began during the LBA following a long separation since the Eneolithic. Although ARS026 exhibits substantial WSH ancestry, strontium isotopic values obtained from his M3 enamel resemble local fauna and fall within the range of the main Khövsgöl cluster (SI Appendix, Fig. S15 and Table S9); however, because the enamel this individual also exhibited elevated manganese levels, postmortem trace element alteration from soil could not be excluded.
Dairy Subsistence and Lactase Persistence.
Contemporary Mongolia has a dairy- and meat-based subsistence economy, and to more precisely understand the role of dairy products in the diets of present-day mobile pastoralists in Khövsgöl aimag, we conducted a detailed nutritional investigation of summer and winter diets. We find that dairy-based foods contribute a mean of 35% total dietary energy, 36–40% total carbohydrate, 24–31% total protein, and 39–40% total fat to rural summer diets in Khövsgöl aimag, with liquid milk and dairy product consumption of 216–283 and 172–198 g/d, respectively (SI Appendix, Table S10 and Dataset S2).
Despite the importance of dairying today, its origins in Mongolia are poorly understood. Given the limited WSH ancestry of the main Khövsgöl cluster, we sought to determine if dairy pastoralism was practiced by this putatively pastoralist LBA population by testing for the presence of milk proteins (30) in the dental calculus of these individuals. We extracted proteins from 12 dental calculus samples representing 9 individuals (SI Appendix, Table S11) and analyzed tryptic peptides using LC-MS/MS (31). Observed modifications included deamidation (N, Q) and oxidation (P, M) (SI Appendix, Table S12). All protein identifications were supported by a minimum of two peptides across the dataset, and only peptides with an E value ≤ 0.001 were assigned; the estimated peptide false-discovery rate (FDR) across the full dataset was 1.0%, and protein FDR was 4.6%. Milk proteins were detected in seven of the nine individuals analyzed (SI Appendix, Table S13 and Dataset S3), confirming that dairy foods were consumed as early as 1456 BCE (1606–1298 BCE, 95% probability of the earliest directly dated individual) (SI Appendix, Fig. S4 and Table S2). Specifically, we detected the milk whey protein β-lactoglobulin (Fig. 5 A and B) and the curd protein α-S1-casein, with peptides matching specifically to sheep (Ovis), goat (Capra), Caprinae, Bovinae, and a subset of Bovidae (Ovis or Bovinae) (Fig. 5C, SI Appendix, Table S13, and Dataset S3). These peptides exhibited asparagine and glutamine deamidation, as expected for ancient proteins (32), and the frequency and distribution of recovered β-lactoglobulin (Fig. 5B) and α-S1-casein peptides closely matched that empirically observed for modern bovine milk (33), thereby providing additional protein identification support through appropriate proteotypic behavior.

Fig. 5.
Presence of ruminant β-lactoglobulin and α-S1-casein milk protein in LBA Khövsgöl dental calculus. (A) B- and Y-ion series for one of the most frequently observed β-lactoglobulin peptides, TPEVD(D/N/K)EALEKFDK, which contains a genus-specific polymorphic residue: D, Bos; N, Ovis; K, Capra. See SI Appendix, Fig. S16 for peptide and fragment ion error distribution graphs. (B) Alignment of observed peptides to the 178 amino acid β-lactoglobulin protein, with peptide taxonomic source indicated by color. Trypsin cut sites are indicated by gray ticks. The position and empirically determined observation frequency of BLG peptides for bovine milk are shown as a heatmap scaled from least observed peptides (light gray) to most frequently observed peptides (dark red), as reported in the Bovine PeptideAtlas (34). Inset displays a 3D model of the β-lactoglobulin protein with observed peptide positions highlighted in black. (C) Taxonomically assigned β-lactoglobulin (black) and α-S1-casein (gray) peptides presented as scaled pie charts on a cladogram of Mongolian dairy domesticates. Bracketed numbers represent the number of peptides assigned to each node. Ruminant milk proteins were well supported, but no cervid, camelid, or equid milk proteins were identified.
Given the evidence for dairy consumption by the LBA Khövsgöl population, we sought to determine if the dairy-adaptive -13910*T (rs4988235) lactase persistence (LP) allele found today in Western steppe (34) and European (35) populations was present among LBA Khövsgöls dairy herders, and we examined this position in our SNP-enriched dataset. The -13910*T LP allele was not found in the LBA Khövsgöls (SI Appendix, Fig. S17 and Table S14), and additionally all observed flanking sequences in the lactase transcriptional enhancer region contained only ancestral alleles.
Discussion
In this study, we find a clear genetic separation between WSH populations and LBA Mongolians more than a millennium after the arrival of WSH at the furthest edges of the Western steppe and the earliest appearance of the WSH Afanasievo cultural elements east of the Altai-Sayan mountain range. This genetic separation between Western and Eastern steppe populations appears to be maintained with very limited gene flow until the end of the LBA, when admixed populations, such as the Karasuk (1200–800 BCE), first appear in the Altai (1) and we observe the first individual with substantial WSH ancestry in the Khövsgöl population, ARS026, directly dated to 1130–900 BCE. Consistent with these observations, we find that the WSH ancestry introduced during these admixture events is more consistent with MBA and LBA steppe populations, such as the Sintashta (2100–1800 BCE), than with earlier EBA populations, such as the Afanasievo (3300–2500 BCE), who do not seem to have genetically contributed to subsequent populations.
Despite the limited gene flow between the Western and Eastern steppes, dairy pastoralism was nevertheless adopted by local non-WSH populations on the Eastern steppe and established as a subsistence strategy by 1300 BCE. Ruminant milk proteins were identified in the dental calculus of most of the tested LBA Khövsgöl individuals, and all identified milk proteins originated from ruminants, specifically the Western dairy domesticates sheep, goat, and Bovinae. These findings suggest that neighboring WSH populations directly or indirectly introduced dairy pastoralism to local indigenous populations through a process of cultural exchange. Further research on other regional cultures in Mongolia, such as Chemurchek, Hemsteg, and Ulaanzuukh, is needed to determine if this pattern of cultural adoption observed among DSKC sites is broadly shared across other Bronze Age cultures throughout the Eastern steppe.
Bronze Age trade and cultural exchange are difficult to observe on the Eastern steppe, where mobile lifestyles and ephemeral habitation sites combine to make household archaeology highly challenging. Burial mounds are typically the most conspicuous features on the landscape, and thus much of Mongolian archaeology is dominated by mortuary archaeology. However, unlike WSH, whose kurgans typically contain a range of grave goods, many LBA mortuary traditions on the Eastern steppe did not include grave goods of any kind other than ritually deposited animal bones from horse, deer, and bovids. Given that Mongolian archaeological collections are typically dominated by human remains with limited occupational materials, the ability to reconstruct technological exchange, human–animal interaction, and secondary product utilization through the analysis of proteins preserved in dental calculus represents an important advance.
The 3,000-y legacy of dairy pastoralism in Mongolia poses challenging questions to grand narratives of human adaptation and natural selection (36). For example, despite evidence of being under strong natural selection (36), LP was not detected among LBA Khövsgöls, and it remains rare (<5%) in contemporary Mongolia even though levels of fresh and fermented dairy product consumption are high (35). Recent studies in Europe and the Near East have found that dairying preceded LP in these regions by at least 5,000 y, suggesting that LP may be irrelevant to the origins and early history of dairying (36). As a non-LP dairying society with a rich prehistory, Mongolia can serve as a model for understanding how other adaptations, such as cultural practices or microbiome alterations (37), may be involved in enabling the adoption and long-term maintenance of a dairy-based subsistence economy. Early herding groups in Mongolia present a historical counter-example to Europe in which WSH migrations resulted in cultural exchange rather than population replacement, and dairying was maintained for millennia without the introgression or selection of LP alleles.
PNAS November 27, 2018 115 (48) E11248-E11255; first published November 5, 2018