|
|
Post by Admin on Jul 24, 2020 9:19:50 GMT
Results The petrous bone of GD13a yielded sequencing libraries comprising 18.57% alignable human reads that were used to generate 1.39-fold genome coverage. The sequence data showed read lengths and nucleotide misincorporation patterns indicative of post-mortem damage, and had low contamination estimates (<1%, see Supplementary Fig. S3). The mitochondrion of GD13a (91.74X) was assigned to haplogroup X, most likely to the subhaplogroup X2, which has been associated with an early expansion from the Near East13,14 and has been found in early Neolithic samples from Anatolia5, Hungary15 and Germany16. We compared GD13a with a number of other ancient genomes and modern populations6,15,16,17,18,19,20,21,22,23,24,25,26,27, using principal component analysis (PCA)28, ADMIXTURE29 and outgroup f3 statistics30 (Fig. 1). GD13a did not cluster with any other early Neolithic individual from Eurasia in any of the analyses. ADMIXTURE and outgroup f3 statistics identified Caucasus Hunter-Gatherers of Western Georgia, just north of the Zagros mountains, as the group genetically most similar to GD13a (Fig. 1B,C), whilst PCA also revealed some affinity with modern Central South Asian populations such as Balochi, Makrani and Brahui (Fig. 1A and Fig. S4). Also genetically close to GD13a were ancient samples from Steppe populations (Yamanya & Afanasievo) that were part of one or more Bronze age migrations into Europe, as well as early Bronze age cultures in that continent (Corded Ware)16,21, in line with previous relationships observed for the Caucasus Hunter-Gatherers24. Figure 1: GD13a appears to be related to Caucasus Hunter Gatherers and to modern South Asian populations. 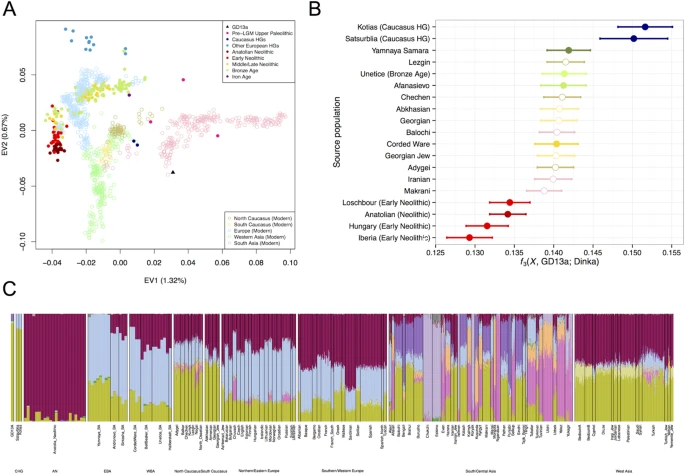 (A) PCA loaded on modern populations (represented by open symbols). Ancient individuals (solid symbols) are projected onto these axes. (B) Outgroup f3( X , GD13a; Dinka), where Caucasus Hunter Gatherers (Kotias and Satsurblia) share the most drift with GD13a. Ancient samples have filled circles whereas modern populations are represented by empty symbols. (C) ADMIXTURE using K = 17, where GD13a appears very similar to Caucasus Hunter Gatherers, and to a lesser extent to modern south Asian populations. We further investigated the relationship between GD13a and Caucasus Hunter-Gatherers using D-statistics30,31 to test whether they formed a clade to the exclusion of other ancient and modern samples (Table S4). A large number of Western Eurasian samples (both modern and ancient) showed significant excess genetic affinity to the Caucasus Hunter-Gatherers, whilst none did with GD13a. Overall, these results point to GD13a having little direct genetic input into later European populations compared to its northern neighbours. To better understand the history of the population to which GD13a belonged, we investigated the distribution of lengths of runs of homozygosity (ROH) (Fig. 2A). A bias towards a high frequency of both long and short ROH is indicative of past strong bottlenecks followed by population re-expansion. GD13a has a distribution with few long ROH (>2 Mb), similar to that of the descendants of Anatolian early farmers (represented by the European farmers NE115 and Stuttgart17). In contrast, both Western17 and Caucasus Hunter-Gatherers24 have relatively more long as well as short ROH. Thus, GD13a is the descendant of a group that had relatively stable demography and did not suffer the bottlenecks that affected more northern populations. Figure 2: GD13a did not undergo a recent large population bottleneck. 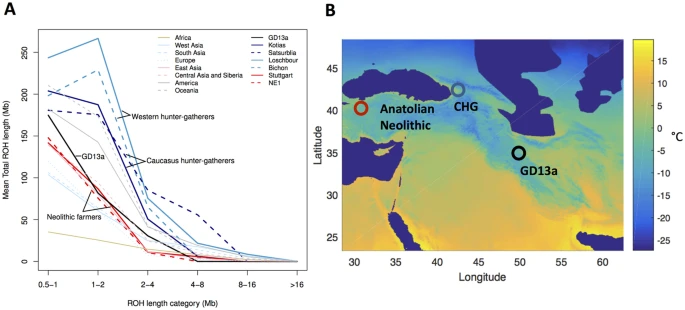 (A) GD13a has similar runs of homozygosity (ROH) lengths to Neolithic individuals, while Caucasus Hunter Gatherers (Kotias and Satsurblia), like European Hunter Gatherers (Loschbour and Bichon), underwent recent large population bottlenecks potentially associated withthe LGM. (B) Map showing geographical location of Anatolian Neolithic samples, Caucasus Hunter Gatherers (CHG) and GD13a. Background colours indicate mean temperature (°C) of coldest quarter during the LGM (data from the worldclim database60 generated by the CCSM4 model)60, with LGM sea levels. Map of populations was generated with MATLAB R2015b (Mathworks, www.mathworks.com/)61. The phenotypic attributes of GD13a are similar to the neighbouring Anatolian early farmers and Caucasus Hunter-Gatherers. Based on diagnostic SNPs, she had dark, black hair and brown eyes (see Supplementary). She lacked the derived variant (rs16891982) of the SLC45A2 gene associated with light skin pigmentation but likely had at least one copy of the derived SLC24A5 allele (rs1426654) associated with the same trait. The derived SLC24A5 variant has been found in both Neolithic farmer and Caucasus Hunter-Gatherer groups5,15,24 suggesting that it was already at appreciable frequency before these populations diverged. Finally, she did not have the most common European variant of the LCT gene (rs4988235) associated with the ability to digest raw milk, consistent with the later emergence of this adaptation5,15,21. It is possible that farmers related to GD13a contributed to the eastern diffusion of agriculture from the Near East that reached Turkmenistan32 by the 6th millennium BP, and continued further east to the Indus Valley33. However, detecting such a contribution is complicated by a later influx from Steppe populations with Caucasus Hunter-Gatherer ancestry during the Bronze Age. We tested whether the Western Eurasian component found in Indian populations can be better attributed to either of these two sources, GD13a and Kotias (a Caucasus Hunter Gatherer), using D-statistics to detect gene flow into an ancestral Indian component (represented by the Onge). Overall, for all tests where a difference could be detected, Kotias and GD13a were equally likely sources (Fig. S9 and Table S6). Whilst the attribution of part of the Western Eurasia component seen in India to Bronze Age migrations is supported by dating of last contact based on patterns of Linkage Disequilibrium34, our analysis highlights the possibility that part of that component might derive from earlier contact during the eastern diffusion of agriculture. |
|
|
|
Post by Admin on Jul 25, 2020 1:04:04 GMT
Discussion
GD13a had little direct genetic input into later European populations compared to the Caucasus Hunter-Gatherers (its northern neighbours) as demonstrated using D-statistics. This lack of connectivity with neighbouring regions might have arisen early on, since we also find that hunter-gatherers from the Caucasus show higher affinity to Western Hunter-Gatherers and early Anatolian farmers; this result suggests the possibility of gene flow between the former and these two latter groups to the exclusion of GD13a. An alternative, but not mutually exclusive, explanation for this pattern is that GD13a might have received genetic input from a source equally distant from all other European populations, and thus basal to them.
The Last Glacial Maximum (LGM) made entire regions in northern Eurasia uninhabitable, and therefore a number of hunter-gatherer populations likely moved to the south. For Europe there may be a separation between Western and Eastern populations with minimal occupation of the Central European plains22. For Eastern Europe, Central Asia and the northern Near East, glaciation itself was less a factor. In these areas, our understanding of how populations weathered the LGM is still vague at best. It has previously been suggested that differences in the frequency of long and short runs of homozygosity in ancient samples may be associated with different demographic experiences during the LGM15,24. Neolithic farmers, with their lower frequency of short ROH, have been argued to have been relatively little affected by the LGM compared to Western and Caucasus Hunter-Gatherers15,24 which are characterised by more long ROH (>2 Mb). GD13a has a profile similar to that of the descendants of Anatolian farmers (i.e. early European farmers), suggesting that her ancestors also faced more benign conditions compared to populations further north. Superimposing the sampling locations of these groups onto climatic reconstructions from the LGM (Fig. 2B), however, does not reveal clear climatic differences among the regions. It is possible that the ancestors of the Anatolian and Ganj Dareh farmers spent the LGM in areas further south or east, which experienced milder climate. But it is also possible that they exploited local pockets of favourable climate (refugia). Whilst high elevation sites in the Zagros were abandoned during the LGM35, there are a number of sites in the valleys that were occupied during that period and might have experienced more favourable conditions36.
The archaeological record indicates an eastward Neolithic expansion from the eastern regions of the Near East into Central and South Asia32,37. Our analysis shows that both the Caucasus Hunter Gatherer Kotias and GD13a are plausible sources for the Eurasian Ancestry found in that part of Asia. Even though part of the Western Eurasian component found in India can be linked to Bronze Age migrations by dating the last contact using Linkage Disequilibrium (thus coming from the Kotias lineage), our results highlight the possibility of an older contribution from a source genetically close to GD13a (which would be hard to disentangle from the later gene flow from the Steppe). Eventually, ancient DNA from the Indus Valley will be needed to detect conclusively whether any genetic traces were left by the eastward Neolithic expansion from the Near East, or whether this process was mostly cultural.
The presence of two distinct lineages (Anatolian-like agriculturalists and Zagros mountain herders) in the Near East at the beginning of the Neolithic transition raises an interesting question regarding the independence of innovations arising at different locations. Even within the Central Zagros, economies vary greatly in their rate and pathway towards Neolithisation35. Ganj Dareh, in the high Zagros, has the earliest known evidence for goat domestication7,8,9, and the foothills of the Zagros mountains have also been argued to have been the site of early farming3. In addition, early sites such as Sheikh-e Abad (11.650-9,600 cal BP) provide evidence of early stages of barley cultivation38. Were these innovations independent of similar achievements that made up the Neolithic package that North West Anatolians brought into Europe? Or were they exchanged culturally? If the latter, it would imply a cultural diffusion in the absence of much genetic interchange.
|
|
|
|
Post by Admin on Jul 26, 2020 3:24:49 GMT
The Formation of Human Populations in South and Central Asia Abstract By sequencing 523 ancient humans, we show that the primary source of ancestry in modern South Asians is a prehistoric genetic gradient between people related to early hunter-gatherers of Iran and southeast Asia. Following the Indus Valley Civilization’s decline, they mixed with people in the southeast to form one of the two main ancestral populations of South Asia whose direct descendants live in southern India. Simultaneously, they mixed with descendants of Steppe pastoralists who spread via Central Asia after 4000 years ago to form the other main ancestral population. The Steppe ancestry in South Asia has the same profile as that in Bronze Age Eastern Europe, tracking a movement of people that affected both regions and that likely spread the unique shared features shared between Indo-Iranian and Balto-Slavic languages. Graphical Abstract  The Bronze Age spread of Yamnaya steppe pastoralist ancestry into two subcontinents, Europe and South Asia. Pie charts reflect the proportion of Yamnaya ancestry, and dates reflect the earliest available ancient DNA with Yamnaya ancestry in each region. There is no ancient DNA yet for the ANI and ASI, so for these the range is inferred statistically. One Sentence Summary: Genome wide ancient DNA from 523 ancient individuals sheds light on genetic exchanges between the Steppe, Iran and South Asia, and highlights the parallel demographic histories of two subcontinents: Europe and South Asia. One Page Summary Introduction and Rationale: To elucidate the extent to which the major cultural transformations of farming, pastoralism and shifts in the distribution of languages in Eurasia were accompanied by movement of people, we report genome-wide ancient DNA data from 523 individuals spanning the last 8000 years mostly from Central Asia and northernmost South Asia. Results: Movements of people following the advent of farming resulted in genetic gradients across Eurasia that can be modeled as mixtures of seven deeply divergent populations. A key gradient formed in southwestern Asia beginning in the Neolithic and continuing into the Bronze Age, with more Anatolian farmer-related ancestry in the west and more Iranian farmer-related ancestry in the east. This cline extended to the desert oases of Central Asia and was the primary source of ancestry in peoples of the Bronze Age Bactria Margiana Archaeological Complex (BMAC). This supports the idea that the archaeologically documented dispersal of domesticates was accompanied by the spread of people from multiple centers of domestication. The main population of the BMAC carried no ancestry from Steppe pastoralists and did not contribute substantially to later South Asians. However, Steppe pastoralist ancestry appeared in outlier individuals at BMAC sites by the turn of the second millennium BCE around the same time as it appeared on the southern Steppe. Using data from ancient individuals from the Swat Valley of northernmost South Asia, we show that Steppe ancestry then integrated further south in the first half of the second millennium BCE, contributing up to 30% of the ancestry of modern groups in South Asia. The Steppe ancestry in South Asia has the same profile as that in Bronze Age Eastern Europe, tracking a movement of people that affected both regions and that likely spread the unique shared features shared between Indo-Iranian and Balto-Slavic languages. The primary ancestral population of modern South Asians is a mixture of people related to early Holocene populations of Iran and South Asia that we detect in outlier individuals from two sites in cultural contact with the Indus Valley Civilization (IVC), making it plausible that it was characteristic of the IVC. After the IVC’s decline, this population mixed with northwestern groups with Steppe ancestry to form the “Ancestral North Indians” (ANI) and with southeastern groups to form the “Ancestral South Indians” (ASI) whose direct descendants live today in tribal groups in southern India. Mixtures of these two post-IVC groups--the ANI and ASI--drive the main gradient of genetic variation in South Asia today. Conclusion: Earlier work recorded massive population movement from the Steppe into Europe early in the 3rd millennium BCE, likely spreading Indo-European languages. We reveal a parallel series of events leading to the spread of Steppe ancestry to South Asia, thereby documenting movements of people that were likely conduits for the spread of Indo-European languages. |
|
|
|
Post by Admin on Jul 27, 2020 1:47:22 GMT
Introduction The past ten thousand years have witnessed profound economic changes driven by the transition from foraging to food production, and have also witnessed dramatic changes in cultural practice evident from archaeology, the distribution of languages, and the written record. The extent to which these changes were associated with movements of people has been a mystery in Central Asia and South Asia in part because of a paucity of ancient DNA. We report genome-wide data from 523 individuals from Central Asia and northernmost South Asia from the Mesolithic period onward (1), and co-analyze them with previously published ancient DNA from across Eurasia and with data from diverse present-day people. In Central Asia, we studied the extent to which the spread of farming and herding practices from the Iranian plateau to the desert oases south of the Steppe was accompanied by movements of people or adoption of ideas from neighboring groups (2–4). For the urban communities of the Bactria Margiana Archaeological Complex (BMAC) in the Bronze Age, we assessed whether the people buried in its cemeteries descended directly from earlier smaller scale food producers, and also documented their genetic heterogeneity (5). Further to the north and east, we showed that the Early Bronze Age spreads of crops and domesticated animals between southwest Asia and eastern Eurasia along the Inner Asian Mountain Corridor (6) was accompanied by movements of people. Finally, we examined when descendants of the Yamnaya, who spread across the Eurasian Steppe beginning around 3300 BCE (7–9), began to appear in Central Asia south of the Steppe. In northernmost South Asia, we report a time transect of more than one hundred individuals beginning ~1200 BCE, which we co-analyze along with modern data from hundreds of present-day South Asian groups, as well as ancient DNA from neighboring regions (10). Previous analyses place the majority of present-day South Asians along a genetic cline (11) that can be modeled as having arisen from mixture of two highly divergent populations after 4000 years ago: the Ancestral North Indians (ANI) who harbor large proportions of ancestry related to West Eurasians, and the Ancestral South Indians (ASI) who are much less closely related to West Eurasians (12). We leveraged ancient DNA to place constraints on the genetic structure of the ANI and ASI and, in conjunction with other lines of evidence, to make inferences about when and where they formed. By modeling modern South Asians along with ancient individuals from sites in cultural contact with the IVC, we inferred a likely genetic signature for people of the Indus Valley Civilization (IVC) which reached its maturity in northwestern South Asia 2600–1900 CE. We also examined when Steppe pastoralist-derived ancestry (9) mixed with groups in South Asia, and placed constraints on whether Steppe-related ancestry or Iranian-related ancestry is more plausibly associated with the spread of Indo-European languages in South Asia. Dataset and Analysis Strategy We generated whole-genome ancient DNA data from 523 previously unsampled ancient individuals and increased the quality of data from 19 previously sequenced individuals. The individuals derive from three broad geographical regions: 182 from Iran and the southern part of Central Asia that we call Turan (present-day Turkmenistan, Uzbekistan, Tajikistan, Afghanistan and Kyrgyzstan), 209 from the Steppe and northern forest zone mostly in present-day Kazakhstan and Russia, and 132 from northern Pakistan. The ancient individuals are from 1) Mesolithic, Copper, Bronze and Iron Age Iran and Turan (12000–1 BCE from 19 sites) including the Bactria Margiana Archaeological Complex (BMAC); 2) early ceramic-using hunter-gatherers from the western Siberian forest zone who we show represent a point along an early Holocene cline of North Eurasians and who emerge as a valuable source population for modeling the ancestry of Central and South Asians (6400–3900 BCE from 2 sites); 3) Copper Age and Bronze Age pastoralists from the central Steppe, including from Bronze Age Kazakhstan (3400–800 BCE from 56 sites); and 4) northernmost South Asia, specifically Late Bronze Age, Iron Age and historical settlements in the Swat and Chitral districts of present-day Pakistan (~1200 BCE - 1700 CE from 12 sites) (Fig. 1, Table S1, (1, 13)). We prepared samples in dedicated clean rooms, extracted DNA (14, 15), and constructed libraries for Illumina sequencing (16, 17). We enriched the libraries for DNA overlapping around 1.2 million single nucleotide polymorphisms (SNPs) (7, 18, 19), sequenced the products on Illumina instruments, and performed quality control (Table S2) (7, 19, 20). Our final dataset after merging with previously reported data (7–9, 16, 18, 19, 21–31) spans 837 ancient individuals that passed all our analysis filters, which included removing individuals determined genetically to be first-degree relatives of other higher coverage individuals (Table S3), and restricting to the 92% of individuals (Table S1) that were represented by at least 15,000 of the targeted SNPs which we found was the number at which we began to be able to reliably estimate proportions of the deeply divergent ancestry sources. The median number of SNPs analyzed per individual was 617,000. We also merged with previously reported whole genome sequencing data from 686 present-day individuals (Table S1), and co-analyzed with 1,789 present-day people from 246 ethnographically-distinct groups in South Asia genotyped at ~600,000 SNPs (Table S5; (13)) (10, 32, 33).  Fig. 1 Overview of ancient DNA data. (A) Distribution of sites and associated archeological or radiocarbon dates along with the number of individuals meeting our analysis thresholds from each site. (B) Locations of ancient individuals for whom we generated ancient DNA that passed our analysis thresholds along with the locations of individuals from 140 groups from present-day South Asia that we analyzed as forming the Modern Indian Cline. Shapes distinguish the individuals from different sites. Data from 106 South Asian groups that do not fit along the Modern Indian Cline as well as AHG are not shown. (C) PCA analysis of ancient and modern individuals projected onto a basis formed by 1,340 present day Eurasians reflects clustering of individuals that mirrors their geographical relationships. An interactive version of this figure is presented in the Online Data Visualizer. We grouped individuals based on archaeological and chronological information, taking advantage of 269 direct radiocarbon dates generated on skeletal material from the individuals from whom we report DNA (Table S4). We further clustered individuals that were genetically indistinguishable within these groupings, and labeled outliers with ancestry that was significantly different from others at the same site and time period (13). For our primary analyses, we did not include individuals that were the sole representatives of their ancestry profiles, thereby reducing the chance that our conclusions were being driven by single individuals with contaminated DNA or misattributed archaeological context. This also ensured that each major analysis grouping was represented by many more SNPs that our minimum cutoff of 15,000 per individual. Thus, all but one analysis cluster included at least one individual covered by >200,000 SNPs, sufficient to support high resolution analysis of population history (19) (the exception is a pair of genetically similar outliers from the site of Gonur who are not the focus of any main analyses). We use Italic font to refer to genetic groupings and plain font to indicate archaeological cultures or sites. To make inferences about population structure, we began by carrying out principal component analysis (PCA) projecting ancient individuals onto the patterns of genetic variation in present-day Eurasians, a procedure that allowed us to obtain meaningful constraints on ancestry even of ancient individuals with limited coverage because each SNP from each individual can be compared to a large reference data set (34–36). This revealed three major clusters strongly correlating to the geographic regions of the Forest Zone/Steppe, Iran/Turan, and South Asia (Fig. 1), a pattern we replicate in ADMIXTURE unsupervised clustering (37). To test if groups of ancient individuals were heterogeneous in their ancestry, we used f4-statistics to measure whether different partitions of these groups into two subgroups differed in their degree of allele sharing to a third group (using a distantly related outgroup as a baseline). We also used f3-statistics to test for admixture (33). To model the ancestry of each group, we used qpAdm, which evaluates whether a tested group is consistent with deriving from a pre-specified number of source populations (relative to a set of outgroups), and if so estimates proportions of ancestry (7). We first used qpAdm to attempt to model groups from the Copper Age and afterward as a mixture of seven “distal” sources related deeply to pre-Copper Age or distantly related modern populations for which we have data (Box 1). In this paper we use the term ‘farmers’ to refer to people who either cultivated crops, or herded animals, or both; this definition covers not only large settled communities but also smaller and probably less sedentary communities like the early herders of the Zagros Mountains of western Iran from the site of Ganj Dareh. The latter kept domesticated animals but did not cultivate crops, and are a key reference population for this study as they had a distinctive ancestry profile that spread widely after the Neolithic (9, 24, 38). We also identified “proximal” models for each group as mixtures of temporally preceding groups (10). We implemented an algorithm, DATES, for estimating the age of population mixtures by measuring the average size of segments of ancestry derive from the admixing populations, an approach whose reliability we verified by computer simulation (10) and that is an improvement relative to methods not optimized for analysis of ancient DNA (33, 39) (the approach’s robustness reflects the fact that it relies for its molecular clock on the accurately measured rate of meiotic recombination in humans (40)). In Box 2, we summarize the findings of these analyses (we use the same headings in Box 2 and the main text to allow cross-referencing), while the Online Data Visualizer (1) allows an interactive exploration of the data. |
|
|
|
Post by Admin on Jul 27, 2020 7:13:04 GMT
Box 1.
Seven Source Populations Used for Distal Ancestry Modeling
Box 2.
Summary of Key Findings
Iran and Turan
A West-to-East Cline of Decreasing Anatolian Farmer-Related Ancestry. There was a west-to-east gradient of ancestry across Eurasia in the Copper Age and Bronze Ages—the Southwest Asian Cline—with more Anatolian farmer-related ancestry in the west and more WSHG- or AASI-related ancestry in the east, superimposed on primary ancestry related to early Iranian farmers. The establishment of this gradient correlates in time to the spread of plant-based agriculture across this region, raising the possibility that people of Anatolian ancestry spread this technology east just as they helped spread it west into Europe.
People of the BMAC Were Not a Major Source of Ancestry for South Asians. The primary BMAC population largely derived from preceding local Copper Age peoples who were in turn closely related to people from the Iranian plateau, and had little of the Steppe ancestry that is ubiquitous in South Asia today.
Steppe Pastoralist-Derived Ancestry Arrived in Turan by 2100 BCE. We find no evidence of Steppe pastoralist-derived ancestry in groups at BMAC sites prior to 2100 BCE, but multiple outlier individuals buried at these sites show that by ~2100–1700 BCE, BMAC communities were regularly interacted with peoples carrying such ancestry.
An Ancestry Profile Widespread During the Indus Valley Civilization. We document a distinctive ancestry profile—45–82% Iranian farmer-related and 11–50% AASI (with negligible Anatolian farmer-related admixture)—that was present at two sites in cultural contact with the Indus Valley Culture (IVC). Combined with our detection of this same ancestry profile (in mixed form) about a millennium later in the post-IVC Swat Valley, this documents an Indus Periphery Cline during the flourishing of the IVC. Ancestors of this group formed by admixture ~5400–3700 BCE. There is little if any Anatolian farmer-related ancestry in the Indus Periphery Cline.
The Steppe and Forest Zone
Ancestry Clines in North Eurasia Established After the Advent of Farming. Prior to the spread of farmers and herders, northern Eurasia was characterized by a west-to-east gradient of very divergent hunter-gatherer populations with increasing proportions of relatedness to present-day East Asians: from Western European Hunter-Gatherers (WEHG), to Eastern European Hunter-Gatherers (EEHG), to West Siberian Hunter-Gatherers (WSHG), to Eastern Siberian Hunter-Gatherers (ESHG). Mixture of people along this ancestry gradient and its counterpart to the south formed five later clines following the advent of farming, the three northern ones of which are the European Cline, the Caucasus Cline, and the Central Asian Cline.
A Distinctive Ancestry Profile Stretching from Eastern Europe to Kazakhstan in the Bronze Age. We add more than one hundred samples from the previously described Western_Steppe_MLBA genetic cluster, including individuals associated with the Corded Ware, Srubnaya, Petrovka, and Sintashta archaeological complexes, and characterized by a mixture of about two-thirds ancestry related to Yamnaya Steppe pastoralists (from the Caucasus Cline), and European farmers (from the European Cline) suggesting that this population formed at the geographic interface of these two groups in Eastern Europe. Our analysis suggests that in the central Steppe and Minusinsk Basin in the Middle to Late Bronze Age, Western_Steppe_MLBA ancestry mixed with about 9% ancestry from previously established people from the region carrying WSHG-related to form a distinctive Central_Steppe_MLBA cluster that was the primary conduit for spreading Yamnaya Steppe pastoralist-derived ancestry to South Asia.
Bidirectional Mobility Along the Inner Asian Mountain Corridor. Beginning in the 3rd millennium BCE and intensifying in the 2nd millennium BCE, we observe multiple individuals in the Central Steppe who lived along the Inner Asian Mountain Corridor and who harbored admixture from Turan, documenting northward movement into the Steppe in this period. By the end of the 2nd millennium BCE, these people were later joined by numerous outlier individuals with East Asian-related admixture which become ubiquitous in the region by the Iron Age (25, 52). This ancestry is also seen in later groups with known cultural impacts on South Asia including Huns, Kushans and Sakas and is hardly present in the two primary ancestral populations of South Asia, suggesting that the Steppe ancestry widespread in South Asia derived from pre-Iron Age Central Asians.
South Asia
Three Ancestry Clines That Succeeded Each Other in Time in South Asia. We identify a unique trio of source populations that fits geographically and temporally diverse South Asians since the Bronze Age: a mixture of AASI, an Indus Periphery Cline group with predominantly Iranian farmer-related ancestry, and Central_Steppe_MLBA. Two-way clines driven more by populations that were mixtures of these three sources succeeded each other in time: prior to 2000 BCE the Indus Periphery Cline had no detectable Steppe ancestry, beginning after 2000 BCE the Steppe Cline, and finally the Modern Indian Cline.
The ASI and ANI Arose as Indus Periphery Cline People Mixed with Groups to the North and East. An ancestry gradient of which the Indus Periphery Cline individuals were a part played a pivotal role in the formation of both the two proximal sources of ancestry in South Asia: a minimum of ~55% Indus Periphery Cline ancestry for the ASI and ~70% for the ANI. Today there are groups in South Asia with very similar ancestry to the statistically reconstructed ASI suggesting that they have essentially direct descendants today. Much of the formation of both the ASI and ANI occurred in the 2nd millennium BCE. Thus, the events that formed both the ASI and ANI overlapped the time of the decline of the IVC.
Steppe Ancestry in South Asia is Primarily from Males and Disproportionately High in Brahmins. Most of the Steppe ancestry in South Asia derives from males, pointing to asymmetric social interaction between descendants of Steppe pastoralists and peoples of the Indus Periphery Cline. Groups that view themselves as being of traditionally priestly status, including traditional custodians of liturgical texts in the early Indo-European language Sanskrit, tend (with exceptions) to have more Steppe ancestry than expected based on ANI-ASI mixture, providing an independent line of evidence for a Steppe origin for South Asia’s Indo-European languages.
|
|

