|
|
Post by Admin on Mar 5, 2021 19:22:02 GMT
Neanderthals in the outermost west - Dr Rebecca Wragg Sykes
What can archaeology tell us about the lives and experiences of Neanderthals in Britain? Dr Rebecca Wragg Sykes, University of Liverpool/Université de Bordeaux explains.
|
|
|
|
Post by Admin on Mar 8, 2021 19:34:04 GMT
To be sure, Neandertals were equipped with a well-known language gene. But some studies cast doubt on whether Neandertals’ anatomy was suitable for producing the sounds of speech: their larynx was shaped differently than that of modern humans. Still, it was considered likely that our cousin species could produce a wide variety of sounds. Whether those sounds rose to the level of actual speech remains controversial. A team led by Mercedes Conde-Valverde of the University of Alcalá in Madrid has now approached the problem from a different angle: The researchers tried to find out whether Neandertals could hear complex language content. If they did have that ability, then it might stand to reason that they could have used it to good effect in listening to spoken speech. Conde-Valverde and her collaborators proceeded by examining the acoustics inside a typical Neandertal inner ear. They recently presented their results in the journal Nature Ecology & Evolution: www.nature.com/articles/s41559-021-01391-6In chimpanzees and various other human ancestors such as Australopithecus africanus and Paranthropus robustus, the inner ear is not designed to resolve the frequency band of language particularly well. The ears of Homo heidelbergensis from the Middle Paleolithic are also not equipped for hearing languages. And what about Neandertals’ ears? Conde-Valverde and her team made 3-D scans using computerized tomography to construct digital models of the structures of the inner ears of five Neandertals. They then compared the sound transmissions within the simulated organs with those of the ears of modern humans, older hominins and chimpanzees. This analysis enabled the Spanish researchers to deduce which pitches each ear was best attuned to. The wider the frequency band that can be detected, the more acoustic signals can be contained within it, enabling the transmission of a distinguishable signal in a short interval. The results from this work is clear: the frequency bandwidth of modern human and Neandertal ears do not differ significantly. Neandertals could therefore hear language just as well as we do—with an acoustic sensorium that is optimally tuned to the frequencies that are particularly relevant for understanding human speech. In contrast to ears of H. heidelbergensis individuals from Spain’s Sima de los Huesos cave, which were analyzed for comparison, Neandertals could hear better at frequencies of four to five kilohertz, an acoustic capacity comparable to that of modern humans. The band of frequencies Neandertals could perceive was also as wide as that of humans. The recent study provides only a hint of whether Neandertals were able to speak. But if they could do so, then they could hear and distinguish between a similarly wide range of sounds as humans. “Neandertals possessed a communication system that was as complex and efficient as modern human speech,” said Conde-Valverde in a recent press release. Her team suggests that Neandertals may have used words with more consonants and significantly fewer vowels than we do today. The researchers’ analyses suggest they could hear subtle differences even between three and five kilohertz. At those ranges, they would be able to hear typical speech sounds such as high-frequency consonants, including so-called voiceless plosives, such as “t” and “k” in the English language, and so-called voiceless fricatives, such as English’s “f,” “s” and “th.” These unvoiced consonant sounds are common in most of our species’ languages. Experts suspect they may be some of the oldest phonemes in human speech. Modern languages also transmit acoustic information in frequency ranges in which so-called formants can be heard, enabling individual vowels to be distinguished at around 2.5 kilohertz. It would be quite unusual if Neandertals could hear consonants better than vowels. By contrast, chimpanzees and many mammals tend to produce vowels in order to communicate. Also, consonant-centered communication would explain why many earlier studies came to the conclusion that Neandertals had no linguistic talent: scientists had often looked for anatomical features that were optimized to produce vowels. “This new study supports a growing consensus that vocal communication was a key part of Neandertals’ everyday life,” says Rebecca Wragg Sykes, an honorary fellow at the University of Liverpool in England and author of Kindred: Neanderthal Life, Love, Death and Art, who was not involved in the work. The ability to communicate, in her view, could have been quite sophisticated. “This new research shows Neandertals and early Homo sapiens likely spoke with broadly similar sounds,” she says, “implying that if attempts were made to talk during interbreeding phases, some common understanding of language [or perhaps an interspecies creole] might have been possible.” |
|
|
|
Post by Admin on Mar 8, 2021 21:01:54 GMT
Neanderthals and Homo sapiens had similar auditory and speech capacities
The study of audition in fossil hominins is of great interest given its relationship with intraspecific vocal communication. While the auditory capacities have been studied in early hominins and in the Middle Pleistocene Sima de los Huesos hominins, less is known about the hearing abilities of the Neanderthals. Here, we provide a detailed approach to their auditory capacities. Relying on computerized tomography scans and a comprehensive model from the field of auditory bioengineering, we have established sound power transmission through the outer and middle ear and calculated the occupied bandwidth in Neanderthals. The occupied bandwidth is directly related to the efficiency of the vocal communication system of a species. Our results show that the occupied bandwidth of Neanderthals was greater than the Sima de los Huesos hominins and similar to extant humans, implying that Neanderthals evolved the auditory capacities to support a vocal communication system as efficient as modern human speech.
The linguistic capacities in Neanderthals have long been an area of active research and debate among palaeoanthropologists, albeit with little resolution1–3. The last two decades have seen increasing archaeological discoveries documenting complex behaviours in Neanderthals. These have been linked to the possible presence of language in Neanderthals, since it seems reasonable to suggest that such behaviours require the presence of a complex and efficient oral communication system3. Nevertheless, a different point of view maintains that the distinctive features of human language, absent in other organisms, include a symbolic element as well as a recursive syntactic process called ‘merge’4. This latter process, at its simplest, uses two syntactic elements and assembles them to form a set and is argued to be exclusive to Homo sapiens and to have appeared no earlier than 100 kyr (refs. 1,4)
Tracing the presence of symbolism and syntactic processes in the course of human evolution currently lies outside the realm of possibility in palaeontology1. Nevertheless, the study of human fossils can prove key to determining whether past human species, and in particular the Neanderthals, possessed the anatomy necessary to produce and perceive an oral communication system as complex and efficient as human speech, the usual (but not exclusive) vehicle for language. In other words, although palaeontology cannot study the evolution of the ‘software’ of language it can contribute to our understanding of the evolution of the ‘hardware’ of speech. In our opinion, to suggest that a past human species, such as the Neanderthals, may have had language, it is not only necessary to establish the presence of symbolism, which can be approached in the archaeological record1 but also to demonstrate the existence of the anatomical bases necessary to produce and perceive articulated speech.
The study of brain endocasts has been one of the classic approaches to interpreting the cognitive and linguistic capacities in Neanderthals. Although the Neanderthals as a species are as enceph-alised as H. sapiens5,6, clear differences have been documented in the cerebral organisation7–9 as well as the allometric trajectories in both phylogenetic10 and ontogenetic11 terms. Nevertheless, the sig-nificance of these differences for inferring cognitive and linguistic capacities in Neanderthals remains actively debated2,4,8,9,12,13.During the last five decades, much debate has centred on whether the supralaryngeal vocal tract in Neanderthals was capable of producing the fundamental sounds of human speech2. While this debate remains unresolved, recent anatomical14–16 and genetic17 data support the idea that Neanderthals could have produced a wide repertoire of acoustic signals, facilitating a complex form of vocal communication. In this context, it would be particularly interesting to establish whether the auditory capacities in Neanderthals were capable, or not, of supporting a vocal communication system as effi-cient as that of our own species.
The auditory capacities in Neanderthals have been indirectly approached through comparative studies of the dimensions and proportions of the ear ossicles18 as well as the functional properties of the middle ear19. These studies found broad similarities between Neanderthals and modern humans in both aspects, suggesting that these similarities might imply similar hearing abilities as well. On the other hand, on the basis of a few features of the external and middle ear, Masali et al.20 proposed that Neanderthals had a slightly higher best audible frequency than modern humans.
More recently, a comprehensive model, based on a large number of anatomical variables directly related to auditory physiology, has been used to estimate the sound power transmission (SPT) through the outer and middle ear in fossil hominins14,21,22. The SPT reflects the acoustic filtering process, leading to a frequency-dependent attenuation of sound power at the cochlear entrance, which largely shapes the resultant audiogram in living subjects23. The SPT directly depends on the dimensions of the different anatomical structures of the outer and middle ear23, making it possible to estimate it in fossil species.
|
|
|
|
Post by Admin on Mar 8, 2021 22:20:59 GMT
At the same time, on the basis of the SPT it is also possible to calculate the occupied bandwidth (OBW), defined as the width of a frequency band such that, below the lower and above the upper frequency limits, the mean powers are each equal to a specified percentage of the total mean power of a given signal spectrum24. In the present study, the OBW includes the range of frequencies which contains at least 90% of the sound power transmitted to the inner ear14,21,22, reflecting the frequency range of maximum auditory sensitivity. The OBW is, in turn, directly related to the bandwidth of the oral communication channel. A wider communication chan-nel bandwidth allows for a larger number of easily distinguishable acoustic signals to be used in the oral communication of a species. This improves the efficiency of communication (that is, the ability to deliver a clear message in the shortest amount of time) since it allows for an increase in the number of phonemes and a reduction of the perception error rate25. The theory of information, as outlined by Shannon25, has been used to estimate the information rate of speech communication26 and forms the basis for recent studies examining the correlation between speech intelligibility and bandwidth for people with normal and impaired hearing27,28. The influence of bandwidth reduction on speech intelligibility is reflected in the definition of the ANSI/ASA S3.5-1997 speech intelligibility index standard29,30. 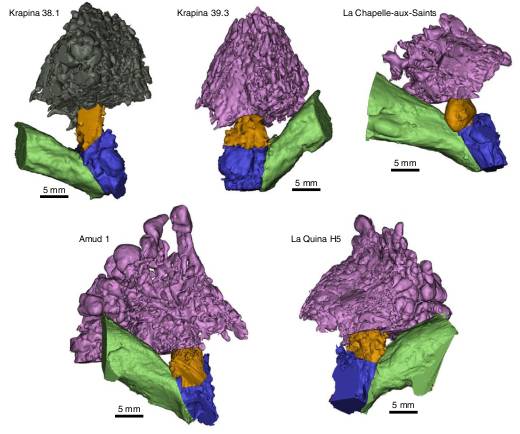 Fig. 1 | Anatomical reconstruction of the external and middle ear cavities in Neanderthals. Thus, there is a clear link between the OBW and the oral communication system in humans. Previous studies carried out on the European Middle Pleistocene fossils from the Sima de los Huesos (SH)14,22 and in the early hominin taxa Australopithecus africanus and Paranthropus robustus from South Africa21 showed that the OBW in the early hominins was quite similar to Pan troglodytes, while the SH fossils had an OBW more similar to, although somewhat narrower than, H. sapiens. In this context, establishing the OBW in Neanderthals would be particularly interesting. If this parameter is similar to that in the SH hominins, it could be argued that the Neanderthals developed more complex behaviours than their ancestors without the need to increase the efficiency of their communication system. Nevertheless, if the OBW in Neanderthals is wider than in the SH hominins, this would show that the increased behavioural complexity of the Neanderthals, compared with their ancestors, was accompanied by an increase in the efficiency of the oral communication system, suggesting a functional link between these two phenomena To establish the SPT and OBW in Neanderthals, we virtually reconstructed the outer and middle ear in five Neanderthal individuals (Fig. 1) from computerized tomography (CT) scans. We have also reconstructed the outer and middle ear in six new SH individuals and have carried out new reconstructions of three previously published individuals14,21,22 on the basis of higher resolution CT scans (Extended Data Fig. 1). These data on the SH hominins are particularly relevant given their close phylogenetic relationship between the SH population and Neanderthals5,31. The fossil samples were compared with a sample of recent H. sapiens (n = 10) published previously21. |
|
|
|
Post by Admin on Mar 9, 2021 3:03:04 GMT
On the basis of anatomical measurements taken on the three-dimensional (3D) models (Extended Data Fig. 2 and Supplementary Table 1), we have calculated the SPT and OBW in the Neanderthals and the expanded SH sample (Table 1, Figs. 2 and 3, Extended Data Fig. 3 and Supplementary Table 2). 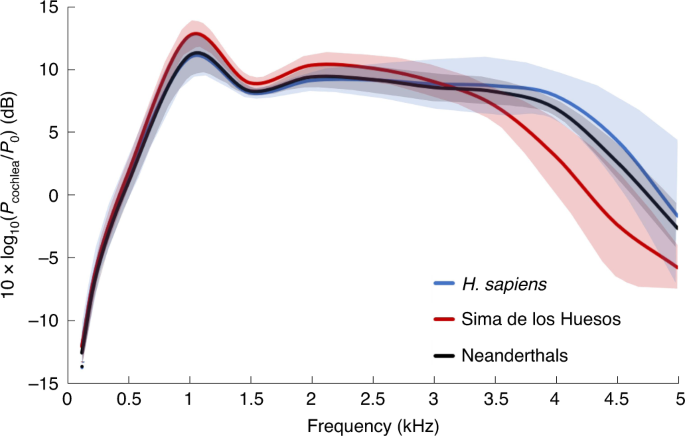 Fig. 2 | SPT in modern humans, the SH hominins and Neanderthals. Results Comparison with previous studies of the SH fossils. The auditory capacities in the SH fossils have been studied previously14,22,32. However, the SH sample used in the present study includes new data in several regards. First, the present study only includes complete individuals, where all the measurements of the outer and middle ear can be taken. Thus, two previously published incomplete individuals (AT-84 and AT-421), which relied on estimating the values for some missing variables22, have been removed from the sample. In addition, previous studies on the auditory capacities of the SH hominins used medical CT scans for SH Crania (Cr.)3, Cr.5 and AT-1907 (ref. 22). These same three individuals have now been CT scanned at a higher resolution, along with all the SH individuals included in this study (Cr.4, Cr.7, Cr.8, Cr.13, Cr.15 and Cr.16) (Supplementary Table 3). As a consequence, we carried out new reconstructions of the external and middle ear cavities of Cr.3, Cr.5 and AT-1907, using the new CT scans and obtained new values for the anatomical variables (Supplementary Table 1). The descriptive statistics of the anatomical variables and the SPT in seven SH individuals, relying on the new reconstructions, were published previously32, but here we include the individual values for these variables in these same individuals, together with the unpublished values in Cr.16 (Supplementary Table 1 for anatomical variables and Supplementary Table 2 for SPT values). In addition, we provide here the individual values for the OBW, a central objective of the present study, for all the SH individuals on the basis of the higher resolution CT scans (Supplementary Table 2). For Cr.3 and AT-1907, the differences between the previ-ous anatomical measurements and the new values are very small (Supplementary Table 1) and the differences for SPT and OBW are also very small (Extended Data Fig. 4). In the case of Cr.5, the dif-ferences between the previously published values and the new data are greater, particularly for the variables LAD, ATM, LEAC and AEAC and the SPT and OBW have also been affected. In particular, the OBW in Cr.5 is narrower than previously published (Extended Data Fig. 4), although, importantly, the new value for OBW falls within the range of variation in the expanded SH sample.The mean value for OBW in the enlarged SH sample is slightly lower than that previously published22. The new SH mean is still significantly higher than the mean OBW in P. troglodytes (Extended Data Fig. 5) but is significantly lower than that in modern humans (Table 1). This lower mean value is mainly due to the low values for OBW obtained in three new individuals included in this study: Cr.4, Cr.8 and Cr.13 (Supplementary Table 2). These are the three largest crania in the SH sample, on the basis of their cranial capaci-ties (Supplementary Table 4). This relationship between cranial size and OBW is explained by the fact that the largest crania also tend to show the longest external auditory canals. Since the length of the external auditory canal is strongly negatively correlated with the OBW (Extended Data Fig. 6), the inclusion of the largest cra-nia in the sample has lowered the mean OBW for the SH homi-nins. Given this relationship between cranial size and the OBW, it is important to emphasise that our Neanderthal comparative sample includes the smallest (La Quina H5) as well as two of the largest (La Chapelle-aux-Saints 1 and Amud 1) known Neanderthal crania (Supplementary Table 2) |
|

