Post by Admin on Mar 9, 2021 4:51:11 GMT
Results in the Neanderthals. The results revealed no statisti-cally significant differences between the Neanderthal and modern human means in any of the anatomical variables (Extended Data Fig. 7) or the SPT (Fig. 2 and Extended Data Fig. 3) or OBW (Table 1 and Fig. 3). Contrary to previous suggestions20, our results for the SPT indicate a similar best frequency in Neanderthals and modern humans. More importantly, the OBW values for Neanderthals fall within the modern human range of variation, except for La Chapelle-aux-Saints 1, which is just below the lower limit of mod-ern humans. At the same time, Neanderthals show some differ-ences from the SH sample in a few anatomical variables (Extended Data Fig. 7), as well as the SPT at frequencies between 4 and 5 kHz (Fig. 2 and Extended Data Fig. 3) and the OBW, which is wider in Neanderthals (Table 1 and Fig. 3) and extended towards higher fre-quencies. These differences in the OBW can be explained by the significantly lower values in Neanderthals than in the SH sample for a few anatomical variables, including the volume of the aditus (VAD), radius of the entrance of the aditus (RAD(entrance)) and length of the external auditory canal (LEAC), which show negative correlations (especially LEAC) with the OBW (Extended Data Fig. 6).
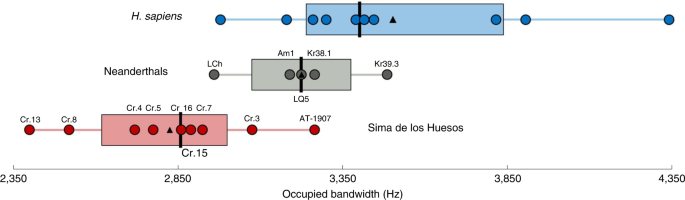
Fig. 3 | OBW in SH, Neanderthals and modern humans
Discussion and conclusions
Our results show that the auditory capacities in Neanderthals do not differ from those in modern humans, including the presence of an extended OBW in both species. Importantly, the OBW in Neanderthals extends towards frequencies that primarily involve consonant production33, suggesting that this may have been an important component of their vocal communication, one that would distinguish them from the pattern of largely vowel-based vocal communication in chimpanzees34 and, indeed, nearly all mammals35. Although much of the acoustic information in human speech is concentrated in the region up to ~2.5 kHz (for example, the first two formant frequencies of the vowels), the region of 3–5 kHz is associated with the production of high-frequency con-sonants33. Consonants in this frequency range mainly consist of the voiceless plosives (stops), such as those associated with the sounds corresponding to the English letters /t/ and /k/, and the voiceless fricatives, including those associated with the sounds correspond-ing to the English letters /f/, /s/ and /th/. While other consonants do occur at lower frequencies, the high-frequency consonants are particularly salient features in human speech. Voiceless plosives are present in over 90% of the world’s languages, with /t/ and /k/ being the most strongly represented36. Similarly, fricative consonants are found in >90% of the world’s languages and among the fricatives, /s/ is the most common voiceless fricative, present in >80% of lan-guages, with /f/ also occurring at high frequencies36. Importantly, because these consonants are voiceless, they do not propagate across the landscape and are limited to short-range intraspecific commu-nication. Indeed, voiceless consonants may represent “…the evolu-tionarily oldest group of consonants”37.In addition, there is evidence that vowels and consonants are processed separately in the human brain38 and that the latter are particularly important for determining word meaning39. In mod-ern human adults with age-related hearing loss, the high-frequency consonants are also the first to be affected, with a concomitant loss in intelligibility and comprehension40, demonstrating a direct link between the OBW and language comprehension. This relation-ship between consonant production and an extended OBW helps make the link between audition and vocalisation explicit and helps explain how an extended OBW corresponds to a vocal communica-tion system in Neanderthals that was as complex and efficient as human speech.Compared to their evolutionary ancestors from the SH, the Neanderthals show an increase in both the OBW and behavioural complexity, including sophisticated subsistence strategies, such as the exploitation of a wide variety of resources41–43, the systematic production and use of fire44,45, the possible construction of musical instruments46 and the expression of symbolic behaviours47–50, includ-ing potential evidence for parietal art51–53. Importantly, this relation-ship between an increase in both behavioural complexity and the OBW was also found in previous studies of the auditory capacities in early hominins (Australopithecus and Paranthropus) and the SH hominins. The early hominins show an OBW that was similar to that in chimpanzees21, while the SH hominins show an OBW that was considerably extended compared with the early hominins, along with an increase in behavioural complexity. This increased behavioural complexity in the SH hominins includes sophisticated stone tool manufacture (mode 2)54, evidence of communal hunt-ing of large game species55, incipient mortuary practices56 and con-specific care57,58. In our opinion, this is strong evidence in favour of the coevolution of increased behavioural complexity and increased efficiency in vocal communication throughout the course of human evolution.
The narrower OBW in the SH hominins, with respect to Neanderthals and modern humans, indicates that the increase in the OBW occurred in both Neanderthals and modern humans after their last common ancestor. This may have evolved through an evolutionary process of adaptive convergence for an increasingly efficient vocal communication system in both lineages. Alternatively, it could be attributed to potential gene flow between the two lineages. We would point out that the Krapina Neanderthals, who show an extended OBW similar to modern humans, date to ~130 thousand years ago (ka)59 and predate the estimated earliest evidence of gene flow between these two species60. This would seem to reject the idea that the Neanderthals acquired their extended OBW through gene flow from modern humans. Nevertheless, it would still be possible that modern humans acquired their extended OBW through gene flow from the Neanderthals. Future study of the auditory capacities in early H. sapiens (older than 100 ka) can reject or reinforce this possibility. If the hypothesis of gene flow is rejected, then the only explanation for the extended OBW in both species would be an adaptive convergence for an increasingly efficient vocal communication system. In summary, our results reinforce the idea that the Neanderthals possessed the same auditory capabilities as H. sapiens necessary to support a vocal communication system as complex and efficient as human speech. It is true that the presence of the anatomical ‘hardware’ necessary to produce human-like speech in the Neanderthals does not necessarily imply the presence of similar mental ‘software’ as in H. sapiens and, by implication, the presence of a language with the same characteristics as that of our own species1. Nevertheless, our results, together with recent discoveries indicating symbolic behaviours in Neanderthals, reinforce the idea that they possessed a type of human language, one that was very different in its complexity and efficiency from any other oral communication system used by non-human organisms on the planet.
Fig. 3 | OBW in SH, Neanderthals and modern humans
Discussion and conclusions
Our results show that the auditory capacities in Neanderthals do not differ from those in modern humans, including the presence of an extended OBW in both species. Importantly, the OBW in Neanderthals extends towards frequencies that primarily involve consonant production33, suggesting that this may have been an important component of their vocal communication, one that would distinguish them from the pattern of largely vowel-based vocal communication in chimpanzees34 and, indeed, nearly all mammals35. Although much of the acoustic information in human speech is concentrated in the region up to ~2.5 kHz (for example, the first two formant frequencies of the vowels), the region of 3–5 kHz is associated with the production of high-frequency con-sonants33. Consonants in this frequency range mainly consist of the voiceless plosives (stops), such as those associated with the sounds corresponding to the English letters /t/ and /k/, and the voiceless fricatives, including those associated with the sounds correspond-ing to the English letters /f/, /s/ and /th/. While other consonants do occur at lower frequencies, the high-frequency consonants are particularly salient features in human speech. Voiceless plosives are present in over 90% of the world’s languages, with /t/ and /k/ being the most strongly represented36. Similarly, fricative consonants are found in >90% of the world’s languages and among the fricatives, /s/ is the most common voiceless fricative, present in >80% of lan-guages, with /f/ also occurring at high frequencies36. Importantly, because these consonants are voiceless, they do not propagate across the landscape and are limited to short-range intraspecific commu-nication. Indeed, voiceless consonants may represent “…the evolu-tionarily oldest group of consonants”37.In addition, there is evidence that vowels and consonants are processed separately in the human brain38 and that the latter are particularly important for determining word meaning39. In mod-ern human adults with age-related hearing loss, the high-frequency consonants are also the first to be affected, with a concomitant loss in intelligibility and comprehension40, demonstrating a direct link between the OBW and language comprehension. This relation-ship between consonant production and an extended OBW helps make the link between audition and vocalisation explicit and helps explain how an extended OBW corresponds to a vocal communica-tion system in Neanderthals that was as complex and efficient as human speech.Compared to their evolutionary ancestors from the SH, the Neanderthals show an increase in both the OBW and behavioural complexity, including sophisticated subsistence strategies, such as the exploitation of a wide variety of resources41–43, the systematic production and use of fire44,45, the possible construction of musical instruments46 and the expression of symbolic behaviours47–50, includ-ing potential evidence for parietal art51–53. Importantly, this relation-ship between an increase in both behavioural complexity and the OBW was also found in previous studies of the auditory capacities in early hominins (Australopithecus and Paranthropus) and the SH hominins. The early hominins show an OBW that was similar to that in chimpanzees21, while the SH hominins show an OBW that was considerably extended compared with the early hominins, along with an increase in behavioural complexity. This increased behavioural complexity in the SH hominins includes sophisticated stone tool manufacture (mode 2)54, evidence of communal hunt-ing of large game species55, incipient mortuary practices56 and con-specific care57,58. In our opinion, this is strong evidence in favour of the coevolution of increased behavioural complexity and increased efficiency in vocal communication throughout the course of human evolution.
The narrower OBW in the SH hominins, with respect to Neanderthals and modern humans, indicates that the increase in the OBW occurred in both Neanderthals and modern humans after their last common ancestor. This may have evolved through an evolutionary process of adaptive convergence for an increasingly efficient vocal communication system in both lineages. Alternatively, it could be attributed to potential gene flow between the two lineages. We would point out that the Krapina Neanderthals, who show an extended OBW similar to modern humans, date to ~130 thousand years ago (ka)59 and predate the estimated earliest evidence of gene flow between these two species60. This would seem to reject the idea that the Neanderthals acquired their extended OBW through gene flow from modern humans. Nevertheless, it would still be possible that modern humans acquired their extended OBW through gene flow from the Neanderthals. Future study of the auditory capacities in early H. sapiens (older than 100 ka) can reject or reinforce this possibility. If the hypothesis of gene flow is rejected, then the only explanation for the extended OBW in both species would be an adaptive convergence for an increasingly efficient vocal communication system. In summary, our results reinforce the idea that the Neanderthals possessed the same auditory capabilities as H. sapiens necessary to support a vocal communication system as complex and efficient as human speech. It is true that the presence of the anatomical ‘hardware’ necessary to produce human-like speech in the Neanderthals does not necessarily imply the presence of similar mental ‘software’ as in H. sapiens and, by implication, the presence of a language with the same characteristics as that of our own species1. Nevertheless, our results, together with recent discoveries indicating symbolic behaviours in Neanderthals, reinforce the idea that they possessed a type of human language, one that was very different in its complexity and efficiency from any other oral communication system used by non-human organisms on the planet.