|
|
Post by Admin on Jun 22, 2021 2:39:35 GMT
The metabolome of humanized Adsl mice We next analyzed the metabolomes of the nine tissues from 9 to 12 adult homozygous humanized mice and their wild-type littermates by gas chromatography coupled with mass spectrometry (GC-MS). We similarly analyzed eight tissues (no testis) from 10 to 12 one-week-old pups (Supplementary file 6). The number of putatively identified metabolites detected varied between 176 and 273 in the adult mice and between 310 and 347 in the young mice. Among those, an average of 280 (median = 273) were detected in at least 50% of the adult and young individuals in each of the nine tissues (Supplementary files 7, 8). A principal component analysis using the mass spectrometric intensities of all metabolites detected revealed one to two outlier samples per tissue (Supplementary file 6). These were excluded from further analyses. Including these samples in the analyses did not qualitatively affect results. Among the organs analyzed, only the brain showed significant differences in metabolite concentrations between the wild type and humanized mice (Figure 5a). To assess the statistical significance, we use a permutation test based on shuffling of sample labels, which estimates a p-value as the ratio of permutations resulting in an equal or greater number of metabolites with significant concentration differences at a nominal t-test significance cutoff of 0.05. According to this test, 36 metabolites showed significant concentration differences in cerebellum of 12-week-old mice (permutation test, p=0.036) and 45 metabolites showed marginally significant differences in the cerebral cortex of 1-week-old mice (permutation test, p=0.081, Supplementary file 9). Thus, the metabolic effects of the two mutations introduced in ADSL are particularly pronounced in the central nervous system. Figure 5 with 1 supplement 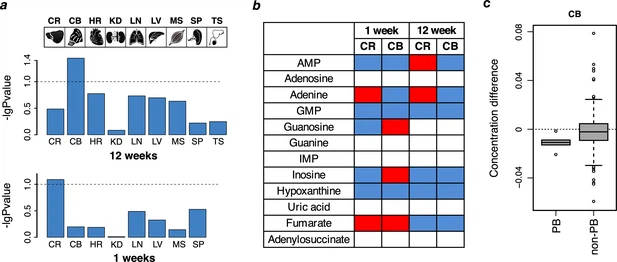 Purine biosynthesis in mice ‘humanized’ for ADSL. (a) Data overview displaying -log10 transformed p-values of the metabolite differences between humanized and wild-type one-week (n = 10–12 per group) and 12-weeks-old (n = 9–12 per group) animals based on all detected metabolites (permutation test based on the Student’s test output). The tissues analyzed are: brain cortex (CR), cerebellum (CB), heart (HR), kidney (KD), lung (LN), liver (LV), muscle (MS), spline (SP), and testis (TS, 12 weeks only). Dotted lines indicate p=0.1. (b) The direction of purine metabolite differences in pairwise comparisons between humanized and wild type mice in the cortex (CR) and cerebellum (CB). Red and blue colors marks higher and lower levels in the humanized mice, respectively, white indicates non-detected metabolites. (c) The distributions of metabolite differences between 12-week-old humanized and wild-type mice for purines biosynthesis (PB) (listed in panel b) and the other metabolites (non-PB) detected using GC-MS in CB of 12-week-old mice (Wilcoxon test, p=0.04). The concentration differences detected in cerebellum of the 12-week-old mice correlated with differences observed in cerebral cortex, even though differences in cortex did not pass the significance cut-off in the permutation test (Pearson correlation, r = 0.84, p<0.0001, n = 29; Figure 5—figure supplement 1a). Similarly, concentration differences detected in cerebral cortex in one-week-old mice correlated with differences observed in cerebellum in the same mice (Pearson correlation, r = 0.77, p<0.0001, n = 45; Figure 5—figure supplement 1b). The concentration differences between wild type and humanized mice furthermore correlated between the 1-week-old and 12-week-old mice both in cortex and in cerebellum (Pearson correlation, r = 0.49 and r = 0.52, p<0.008, n = 28 and n = 31, respectively; Figure 5—figure supplement 1e,f). By contrast, the correlation between metabolic differences detected in brain and the other tissues was weaker (p<10−8 for brain tissues and p>10−4 for other tissues) (p<10−8; Figure 5—figure supplement 1c,d). Thus, in the humanized mouse, the effects of the substitutions in ADSL are seen in the cerebral cortex and cerebellum in both young and adult animals. |
|
|
|
Post by Admin on Jun 22, 2021 5:53:08 GMT
Activity and stability of humanized mouse ADSL To investigate how the humanized form of the mouse ADSL enzyme may influence purine biosynthesis, we synthesized mouse wild-type (wt) ADSL and mouse A429V ADSL and inserted them in expression vectors that include N-terminal polyhistidine tags (pET-14b vector) (Lee and Colman, 2007). We analyzed the conversion of SAICAR to AICAR, and the conversion of SAMP to AMP, in the presence of excess substrate by measuring the rate of production of AICAR and AMP (Figure 6a) and found no differences in the kinetics of either reaction between wt and A429V ADSL (t-test, p>0.05). Figure 6 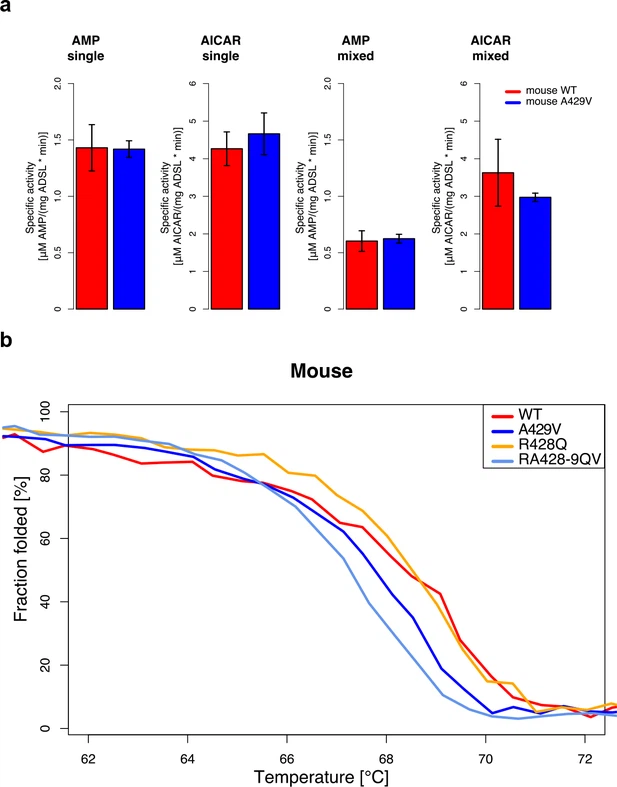 Characterization of mouse ADSL forms. (a) Enzyme kinetics tested in substrate excess for the products (AICAR and AMP, see text above the plots) with one substrate (single) or with both substrates (mixed) in the reaction mix. Specific activities of wild type and humanized A429V versions do not differ (t-test, p>0.05). Error bars represent standard error of the mean from three experiments using two batches of proteins. (b) Protein melting measured by CD at 222 nm for WT, A429V, R428Q, and RA428-9QV ADSL versions. Lines represent the averages over four experiments. The A429V (t-test, p=0.10) and RA428-9QV versions are less stable than the other proteins (t-test, p=0.03, at denaturation midpoint (dm). dm(WT)=68.3 +/- 0.2°C, dm(A429V)=67.9 +/- 0.2°C, dm(R428Q)=68.6 +/- 0.0°C, and dm(RA428-9QV)=67.3 +/- 0.3°C). |
|
|
|
Post by Admin on Jun 22, 2021 19:29:52 GMT
Characterization of human and Neandertal ADSL Previous work has shown that the modern human version of ADSL is less stable than the ancestral, Neandertal-like form in vitro (Van Laer et al., 2018). To ensure that the two forms of the ADSL protein which differ only at position 429 in the protein are both enzymatically active in living cells we show that they both are able to rescue Chinese hamster ovary cells that lack ADSL activity (AdeI cells, Vliet et al., 2011; data not shown). We then expressed the modern human and Neandertal versions of ADSL in E. coli and isolated the two proteins by N-terminal His-tags (Lee and Colman, 2007). In agreement with previous results (Van Laer et al., 2018), circular dichroism spectra of both protein versions exhibited predominantly alpha-helical characteristics (data not shown). To investigate if the enzymatic activities of the human and Neandertal versions of ADSL differ, we tested their ability to convert SAICAR to AICAR, and SAMP to AMP, as described above. No differences in specific activity between the two enzymes were detected using either substrate (SAMP, t-test, p=0.41 or SAICAR, t-test, p=0.81, Figure 7a). Figure 7 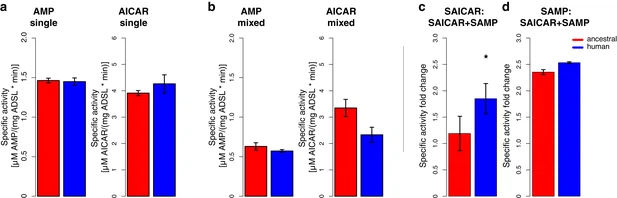 Specific activity and cooperativity of human and ancestral ADSL versions. Specific activity of SAMP to AMP and SAICAR to AICAR catalysis for the Neandertal-like and modern human forms of ADSL when incubated (a) with the required substrate separately (‘single’) (t-test, p>0.05); (b) and with both substrates (SAICAR and SAMP) together (‘mixed’) (t-test, p>0.05). (c, d) The ratios of single substrate to mixed substrate specific activities for the ancestral and human versions of the enzyme. The human version shows a higher reduction in specific activity for SAICAR to AICAR catalysis in the presence of both substrates than the ancestral version (t-test, p=0.01) (c) which is not seen for SAMP to AMP conversion (d) (t-test, p=0.49). Error bars represent standard errors among three experiments. To investigate if cooperativity between the two activities of ADSL may be affected by the A429V substitution, we incubated the two forms of the enzyme with equimolar amounts of SAMP and SAICAR and calculated relative values of ADSL protein activity when presented with a single substrate (Figure 7a) or an equimolar mixture of SAMP and SAICAR (Figure 7b). The conversion of SAMP to AMP is reduced approximately to the same extent (~2.4-fold, t-test, p=0.49) for the modern human and Neandertal-like forms of the enzyme when both substrates are present (Figure 7d). In contrast, the conversion of SAICAR to AICAR is reduced about 1.8-fold for the modern human form in the presence of both substrates (p=0.026) while the ancestral form of the enzyme shows no reduction (p=0.44; Figure 7c). This suggests that the modern human enzyme produces less AICAR than the ancestral enzyme under physiological conditions and may warrant further investigation. Stability of modern human and ancestral ADSL To investigate the effects of the A429V substitution in the human protein, we compared the stability of the secondary structure of the protein as described above. The temperature at which 50% of the protein is inferred to be folded is 67.0°C (± SEM = 0.10°C) for the modern human form of the protein and 68.0°C (± SEM = 0.2°C) for the Neandertal-like form of the protein (Figure 8a). Thus, as previously shown (Van Laer et al., 2018), modern human ADSL is less stable than ancestral ADSL. Figure 8 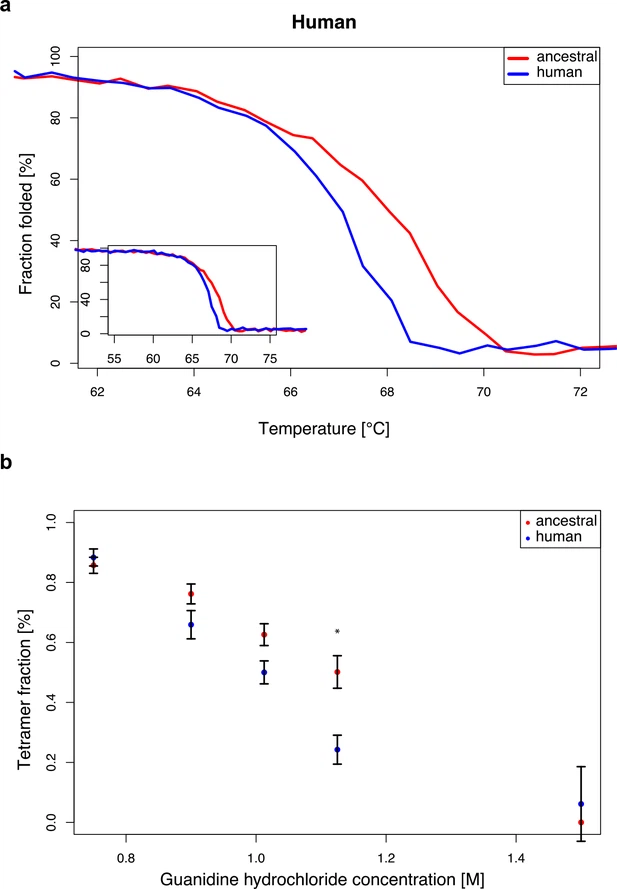 Stability of modern human and Neandertal-like ADSL. To investigate the effect of the difference in secondary structure stability on the stability of ADSL quaternary structures, we exposed the proteins to 0.75M, 0.90M, 1.0125M, 1.125M, and 1.5M guanidine hydrochloride (GdnHCl) overnight at room temperature and analyzed the proteins by native gel electrophoresis. Bands corresponding to the predicted size of the ADSL tetramer (~223 kDa) and monomer (~56 kDa) were observed and the relative proportion of tetrameric protein was estimated. Figure 8b shows that at 1.0125 M GdnHCl about 50% of the modern human version of ADSL was present as tetramers whereas for the ancestral form this was the case at 1.125 M GdnHCl. Thus, both the secondary and quaternary structures of the ancestral form of ADSL are more stable than for the modern human form. |
|
|
|
Post by Admin on Jun 22, 2021 21:08:45 GMT
Ancestralized ADSL in human cells To investigate how the A429V substitution affects the metabolome of living human cells, we used CRISPR-Cas9 to introduce the nucleotide substitution in the ADSL gene that reverts the valine at position 429 to the ancestral alanine residue in human 409B2 cells (Riken BioResource Center). We isolated three independent cell lines and we verified that the intended nucleotide substitution had occurred in each of them by sequencing a segment of the ADSL gene and excluded deletions of the target site by quantitative PCR (data not shown). We also isolated six independent lines that had been subjected to the CRISPR-Cas9 procedure but did not exhibit any mutation in the sequenced DNA segment. We expanded 10 separate cultures of the three edited cell lines and 19 cultures of three control lines (Figure 9—figure supplement 1). Each of these cultures were analyzed by LC-MS in positive and negative modes. A total of 8849 metabolite peaks were detected in positive and negative mode (Supplementary file 10). Among them, 1989 were computationally annotated using KEGG. Strikingly, out of 10 detected metabolites downstream of ADSL, all but one (adenine) have lower concentrations in the wild type cells (binom. test, p=0.01; Wilcoxon test, p=0.06, Figure 9a). These metabolites are present in lower amounts in the wild-type human cells than the cell carrying the V429A substitution in ADSL (Wilcoxon test, p=0.06; Figure 9b). Thus, in human cells, the ancestral form of ADSL supports a higher level of purine biosynthesis than the modern human form. Figure 9 with 1 supplement 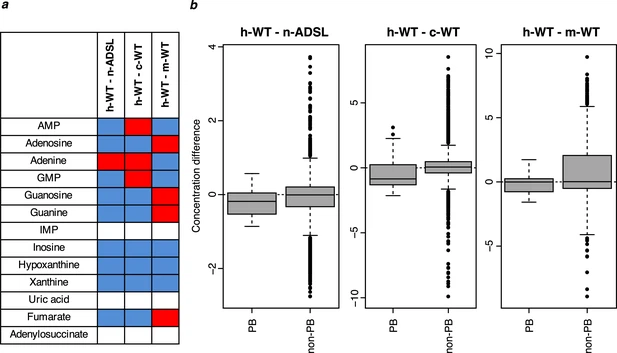 Purine biosynthesis in human cells carrying the ADSL V429A substitution as well as in human, chimpanzee, and macaque wild type cells. (a) The direction of differences in metabolite levels in pairwise comparisons between wild type human cells (h-WT, 19 cultures of three cell lines) and human cells carrying the ADSL V429A substitution (n-ADSL, 10 cultures of three cell lines), chimpanzee cells (c-WT, nine cultures from three cell lines) and macaque cells (m-WT, three cultures from one cell line). Red and blue colors mark higher and lower metabolite levels in human wild-type cells, respectively, white indicates non-detected metabolites. (b) Distributions of differences in metabolite levels between h-WT cells and n-ADSL, c-WT, and m-WT cells, respectively, for purines biosynthesis (PB) metabolites (listed in panel a) and other metabolites (non-PB) Wilcoxon test, p=0.06 (h-ADSL), p=0.01 (c-WT), and p=0.04 (m-WT). Comparison to chimpanzee and macaque cells To compare these results to purine biosynthesis in chimpanzees and macaques, we similarly analyzed nine cultures of three different chimpanzee pluripotent cell lines and three cultures of a macaque pluripotent cell line (Figure 9—figure supplement 1). Out of 1883 metabolite peaks that differ significantly (Wilcoxon test, BH-adjusted p<0.05) between the human and chimpanzee wild-type cells, four correspond to purine biosynthesis metabolites (guanosine, inosine, hypoxanthine, xanthine) and are present at lower levels in human cells compared to chimpanzee cells. The same 10 metabolites as in the comparison to the ancestralized human cells were detected in the chimpanzee cells. Seven of them are present in lower amounts in the human cells (Figure 9a) and as a group they are present in lower concentrations in the human than in the chimpanzee cells when compared to metabolites in other pathways (Wilcox test, p=0.01, Figure 9b). Six of the 10 purine biosynthesis metabolites are present at significantly lower concentrations in humans than in macaques when analyzed individually (Figure 9a) and as a group they are present at lower concentrations than metabolites in other pathways (Wilcoxon test, p=0.04, Figure 9b). Thus, reversal of the A429V substitution in ADSL in human cells results in an increase in purine biosynthesis similar in magnitude to what is observed when chimpanzee cells are compared to human cells. |
|
|
|
Post by Admin on Jun 23, 2021 2:07:53 GMT
The ancestors of modern humans diverged from their closest evolutionary relatives, Neandertals and Denisovans, on the order of 600,000 years ago (Prüfer et al., 2014). About 10 times further back the ancestors of hominins diverged from the ancestors of chimpanzees and bonobos (Langergraber et al., 2012). Whereas some phenotypic differences between present-day humans and apes have been shown to be due to genetic features unique to humans, especially gene duplications (e.g. Charrier et al., 2012; Nuttle et al., 2016; Florio et al., 2018) and changes involving gene expression (reviewed by Doan et al., 2018), no organismal traits that distinguish present-day modern humans from Neandertals and Denisovans have been associated with specific nucleotide changes. Whereas most phenotypic features that distinguish present-day humans from the apes and from their extinct hominin relatives are likely to be genetically complex, metabolic differences may sometimes have a comparatively simple genetic background because genomic changes that affect genes encoding enzymes may affect flux in metabolic pathways.
To find metabolic differences that set humans apart from their closest evolutionary relatives we investigated the metabolomes of the brain, muscle, and kidney in humans, apes, and monkeys. Whereas we find no such differences among the metabolites analyzed in muscle and kidney, in the brain, steady state abundance of many compounds involved in amino acid metabolism are present in higher or lower levels in humans versus other primates and metabolites involved in oxidative phosphorylation are present in lower amounts. In the future, it may be of interest to investigate more samples and the consequences of these human-specific metabolic features. Metabolites in purine biosynthesis stand out in that they are present in lower concentrations in humans than in the other primates analyzed in all tissues analyzed, although most drastically in brain.
Humans and apes diverged so long ago that almost every gene carries changes that may potentially alter its function by affecting its regulation or the structure of the encoded gene product. In contrast, modern humans and Neandertals and Denisovans diverged so recently that for about 90% of the genome, the two archaic human groups fall within the variation of present-day humans (Green et al., 2010). Furthermore, when modern and archaic humans met about 50,000 years ago, they interbred. This resulted in that Neandertal DNA fragments that together make up about half of the Neandertal genome exist in present-day humans (Sankararaman et al., 2014). The number of proteins that carry amino acid substitutions in all or almost all humans that differ from Neandertals and apes is therefore only about one hundred (Pääbo, 2014). It is unclear if any of these substitutions have any functional consequences.
The alanine to valine substitution at position 429 in ADSL is one of this small number of substitutions. Position 429 is conserved as alanine in most tetrapods suggesting that it may be of importance. Position 429 is also located only three positions away from position 426, where an arginine to histidine substitution causes the most common form of adenylosuccinase deficiency in present-day humans (Edery et al., 2003; Kmoch, 2000; Maaswinkel-Mooij et al., 1997; Marie et al., 1999; Race et al., 2000). Further evidence suggesting that a change in ADSL may have been of importance in the evolution of modern humans comes from a screen for genomic regions that have experienced selective sweeps in humans after their split from Neandertals but before the separation of Africans and Eurasians (Racimo, 2016). In that study, a genomic region centered around ADSL is among the top 20 candidate regions, although it also contains other genes. Furthermore, previous work has shown that the A429V substitution reduces the thermal stability of the ADSL protein in vitro (Van Laer et al., 2018). We therefore decided to analyze if it might be involved in the reduced purine biosynthesis seen in present-day humans by investigating the function of the ancestral, Neandertal-like and the derived, modern human-like forms of ADSL in vitro and in vivo.
We confirm the previous finding (Van Laer et al., 2018) that the A429V substitution does not affect the kinetic properties of the ADSL enzyme but decreases its thermal stability and show that the substitution also decreases the stability of the tetrameric complex of the enzyme. When introduced in the mouse ADSL protein in conjunction with a primate-specific substitution at the adjacent position 428, it reduces the enzymatic activity detected in nine tissues analyzed, most drastically in the brain, and results in a reduction in purine biosynthesis, thus recapitulating differences seen between humans and chimpanzees and macaques.
To investigate how the A429V substitution may affect the metabolism of human cells, we used CRISPR-Cas9 to introduce the ancestral, Neandertal-like substitution into human cells. The concentrations of nine out of 10 detected metabolites downstream of ADSL in purine biosynthesis are increased in human cells carrying the ancestral substitution. In chimpanzee and macaque cells, most of the same 10 metabolites are similarly increased resulting in that the pathway as a whole is increased relative to other pathways. Notably, the expression of ADSL messenger RNA does not differ between human and chimpanzee cells, nor between wild type and ancestralized cells (not shown). Thus, the A429V substitution is responsible for much or all of the difference in purine biosynthesis observed when human tissues are compared to ape and monkey tissues, indicating that this change in metabolism occurred in humans after their separation from the ancestor shared with Neandertals and Denisovans.
It is interesting that although ADSL is expressed and functions in all tissues, the down-regulation of purine biosynthesis in humans relative to apes, and in humanized mice relative to wild-type mice, is most pronounced in the brain. It is also interesting that mutations in humans that affect enzymes involved in purine metabolism have more pathological consequences in the nervous system than in other organs (Fumagalli et al., 2017; Micheli et al., 2011). Although no overt morphological or other brain-related phenotypes are observed in the mice humanized at pos. 428 and 429 (data not shown), it is thus possible that the A429V substitution in ADSL has contributed to human-specific changes in brain development and function. Future work will have to address this and other possibilities.
|
|

