Post by Admin on Aug 29, 2020 7:09:15 GMT
Results
We successfully reconstructed complete or almost entire mitochondrial genomes for 26 individuals, 3 from Shekerdja mogila, 1 from Gabrova mogila and 22 from Bereketska mogila (Table 2). All the resulted sequences reach the standard quality requested to guaranty the reliability of the NGS data; CtoT patterns range between 20% to 46%, average fragment size vary from 44.4 base pair (bp) to 67.4 bp and no significant levels of present-day human contamination were detected (Table 2). Only one sample, BM-51, showed a high level of contamination and was not considered in the following statistical analyses.
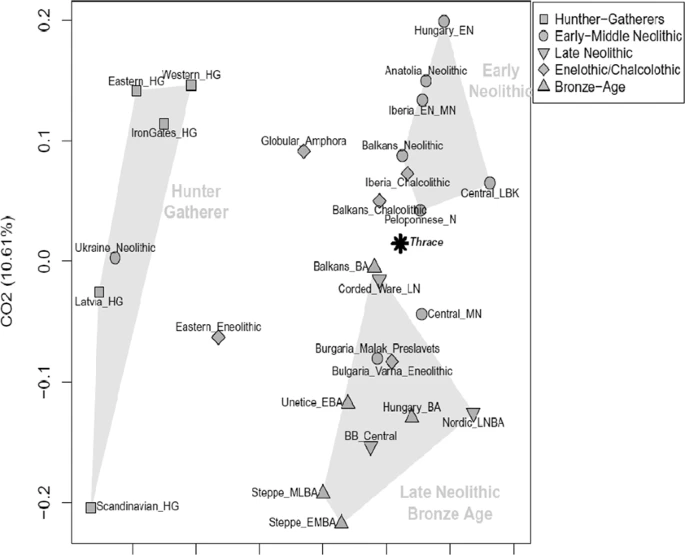
The direct radiocarbon dating performed on the samples BM 44, SM 8.1 and GM 30.3 placed the remains at II-III millennium BC (CEDAD, Centro di DAtazione e Diagnostica, Univerità del Salento, Italy) (Table 1), that corresponds to the age estimated according to the archaeological record. The mtDNA sequences obtained were assigned to 21 different haplogroups, representative of the mitochondrial variability of Western Eurasia (Table 2 and Supplementary Table S1). Phylogenetic links between haplotypes of the Thracian samples and comparison ancient data are shown in the Median Joining Network (Fig. 2). Most of the Thracian individuals belong to sub-lineages of the macro-haplogroup H, which accounts for an overall frequency of 33%. This is the most frequent mitochondrial lineage in present-day Europe, representing over 40% of the total mtDNA variability20. Its frequency observed in the Thracians samples is almost similar to the frequency in contemporary European population. Two individuals belong to haplogroup HV, an ancient European lineage likely originating in the Mediterranean region during the Last Glacial Maximum (LGM)21. In ancient samples, HV has been identified in one Mesolithic specimen from Sicily22 and in early Neolithic remains from Spain23, Germany8 and Russia18,24; Mathieson et al.18 reported a HV haplotype in one sample from Serbia dating from 5800 BCE. Moreover, haplogroup HV was observed in Copper Age specimens from Scotland, Hungary and Germany25 and in Hungarian and Israeli samples from the Chalcolithic period26,27
Figure 2
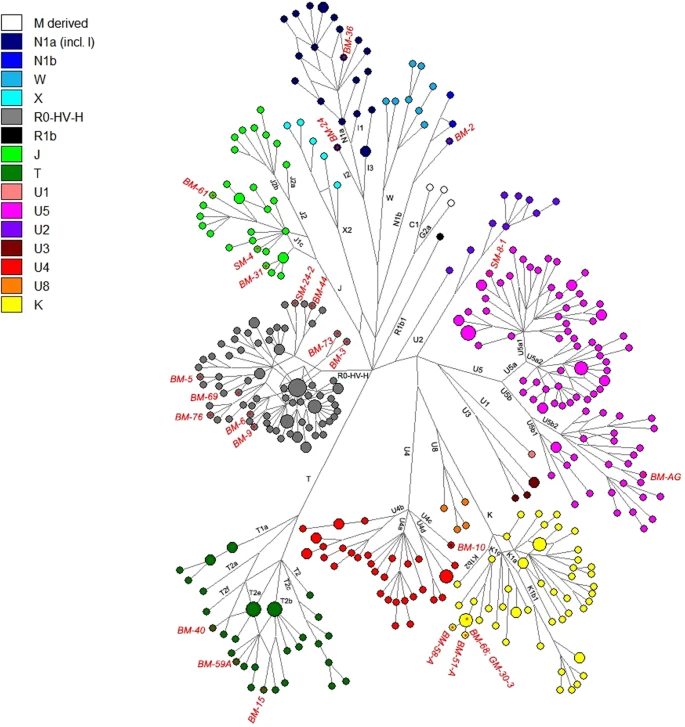
We found four individuals belonging to haplogroup K1c (GM-30.3, BM-51A, BM-58A and BM-68). All the haplotypes contain the expected K1c defining variants with the following private polymorphisms: GM-30.3, 309.1T, 310C, 7441T and 16519C; BM-51A, 16519C; BM-58A, 310C, 513.1CA and 16519C; BM-68, 5297T and 16519C. Nowadays the highest observed European frequency of the lineage K is in Bulgaria (13.3%)28 and K1c is particularly common in Slavic-speaking countries. In ancient populations, the haplogroup K1c has been identified in six hunter-gatherers dated before the arrival of farming (one in Romania, three in Serbia18 and two in Greece29), in two Bronze-Age individuals from Hungary and Bulgaria18,30,31 and in two Central-Europe farmers associated with the Bell-Beaker culture25,32,33. The phylogenetic network analysis (Fig. 2) reveals that the detected K1c haplotypes in Thracians are closely related to hunter-gatherers from Iron Gates and Bronze Age individuals from Bulgaria and Hungary.
Three samples belong to haplogroup J1c (SM-4, BM-31 and BM-61). The SM-4 individual shows three personal transitions previously identified at positions 199C, 8730G and 13928A, and a private mutation at 13686G. The haplotypes of samples BM-31 and BM-61 fall within the sub-haplogropus J1c9 and J1c6, respectively. Currently, J1c, which dates to ∼16 ka ago, is found mainly in Europe, especially in Central Europe, Balkans and Ukraine, where it encompasses almost 80% of total J1 lineages. Pala et al.34 suggested that during the LGM, haplogroup J sub-lineages arose in the Near Eastern refugia and recolonized Europe following the end of the last glaciation. In particular, J1c is not yet found in any hunter-gatherers, and the oldest individuals belonging to this lineage were found in Iran35 and in Anatolia30 dating to 8000-7700 BCE. It is possible that J1c arrived in Thracia from Anatolia during the early stages of the Neolithic expansion. The expansion of farmers played an important role also in the diffusion of haplogroup T, which has been found in three Thracian samples with the T2b (BM-15 and BM-59A) and T2e (BM-40) sub-lineages. Pala et al.34 particularly suggested that these lineages entered Europe from Anatolia in the Late Glacial period, and have been later diffused around Europe by Neolithic agriculturalists after intermingling with the inhabitants of Southeast Europe. Overall, while haplogroups H, K, J and T arose throughout the Neolithic increasing frequencies in different later communities and present-day European populations, the haplogroup U sub-lineages including U2, U4, U5 and U8 instead mark the genetic pool of European pre-LGM hunter-gatherers36,37,38.
We successfully reconstructed complete or almost entire mitochondrial genomes for 26 individuals, 3 from Shekerdja mogila, 1 from Gabrova mogila and 22 from Bereketska mogila (Table 2). All the resulted sequences reach the standard quality requested to guaranty the reliability of the NGS data; CtoT patterns range between 20% to 46%, average fragment size vary from 44.4 base pair (bp) to 67.4 bp and no significant levels of present-day human contamination were detected (Table 2). Only one sample, BM-51, showed a high level of contamination and was not considered in the following statistical analyses.
The direct radiocarbon dating performed on the samples BM 44, SM 8.1 and GM 30.3 placed the remains at II-III millennium BC (CEDAD, Centro di DAtazione e Diagnostica, Univerità del Salento, Italy) (Table 1), that corresponds to the age estimated according to the archaeological record. The mtDNA sequences obtained were assigned to 21 different haplogroups, representative of the mitochondrial variability of Western Eurasia (Table 2 and Supplementary Table S1). Phylogenetic links between haplotypes of the Thracian samples and comparison ancient data are shown in the Median Joining Network (Fig. 2). Most of the Thracian individuals belong to sub-lineages of the macro-haplogroup H, which accounts for an overall frequency of 33%. This is the most frequent mitochondrial lineage in present-day Europe, representing over 40% of the total mtDNA variability20. Its frequency observed in the Thracians samples is almost similar to the frequency in contemporary European population. Two individuals belong to haplogroup HV, an ancient European lineage likely originating in the Mediterranean region during the Last Glacial Maximum (LGM)21. In ancient samples, HV has been identified in one Mesolithic specimen from Sicily22 and in early Neolithic remains from Spain23, Germany8 and Russia18,24; Mathieson et al.18 reported a HV haplotype in one sample from Serbia dating from 5800 BCE. Moreover, haplogroup HV was observed in Copper Age specimens from Scotland, Hungary and Germany25 and in Hungarian and Israeli samples from the Chalcolithic period26,27
Figure 2
We found four individuals belonging to haplogroup K1c (GM-30.3, BM-51A, BM-58A and BM-68). All the haplotypes contain the expected K1c defining variants with the following private polymorphisms: GM-30.3, 309.1T, 310C, 7441T and 16519C; BM-51A, 16519C; BM-58A, 310C, 513.1CA and 16519C; BM-68, 5297T and 16519C. Nowadays the highest observed European frequency of the lineage K is in Bulgaria (13.3%)28 and K1c is particularly common in Slavic-speaking countries. In ancient populations, the haplogroup K1c has been identified in six hunter-gatherers dated before the arrival of farming (one in Romania, three in Serbia18 and two in Greece29), in two Bronze-Age individuals from Hungary and Bulgaria18,30,31 and in two Central-Europe farmers associated with the Bell-Beaker culture25,32,33. The phylogenetic network analysis (Fig. 2) reveals that the detected K1c haplotypes in Thracians are closely related to hunter-gatherers from Iron Gates and Bronze Age individuals from Bulgaria and Hungary.
Three samples belong to haplogroup J1c (SM-4, BM-31 and BM-61). The SM-4 individual shows three personal transitions previously identified at positions 199C, 8730G and 13928A, and a private mutation at 13686G. The haplotypes of samples BM-31 and BM-61 fall within the sub-haplogropus J1c9 and J1c6, respectively. Currently, J1c, which dates to ∼16 ka ago, is found mainly in Europe, especially in Central Europe, Balkans and Ukraine, where it encompasses almost 80% of total J1 lineages. Pala et al.34 suggested that during the LGM, haplogroup J sub-lineages arose in the Near Eastern refugia and recolonized Europe following the end of the last glaciation. In particular, J1c is not yet found in any hunter-gatherers, and the oldest individuals belonging to this lineage were found in Iran35 and in Anatolia30 dating to 8000-7700 BCE. It is possible that J1c arrived in Thracia from Anatolia during the early stages of the Neolithic expansion. The expansion of farmers played an important role also in the diffusion of haplogroup T, which has been found in three Thracian samples with the T2b (BM-15 and BM-59A) and T2e (BM-40) sub-lineages. Pala et al.34 particularly suggested that these lineages entered Europe from Anatolia in the Late Glacial period, and have been later diffused around Europe by Neolithic agriculturalists after intermingling with the inhabitants of Southeast Europe. Overall, while haplogroups H, K, J and T arose throughout the Neolithic increasing frequencies in different later communities and present-day European populations, the haplogroup U sub-lineages including U2, U4, U5 and U8 instead mark the genetic pool of European pre-LGM hunter-gatherers36,37,38.