|
|
Post by Admin on Jun 30, 2021 21:22:30 GMT
Life reconstruction The overall size, robustness, thick and strong supraorbital tori, large mastoid processes, and salient temporal lines of the Harbin cranium suggest that it probably represents a male individual. The ectocranial sutures at the midlambdoid, lambdoid, obelion, anterior sagittal, superior sphenotemporal, incisive, anterior and posterior median palatine, and transverse palatine are all completely obliterated. The ectocranial sutures at bregma, midcoronal, pterion, sphenofrontal, and inferior sphenotemporal show significant closure. For the standard of H. sapiens, the ectocranial suture composite scores would suggest an old adult around 50 years old.11,12 However, the tooth wear seems to suggest a younger age. The only preserved M2 still has much enamel present, and dentine exposure is present on the protocone and paracone. The relatively complete ectocranial suture closure may be related to the robustness of the Harbin cranium. The large square eye sockets with strong supraorbital tori indicate deep eyes. The large and wide piriform aperture indicates a large and bulbous nose.  Figure 3 Life reconstruction of the Harbin cranium The expanded paranasal region and relatively projecting middle face are matched with flat and short modern human-like cheek regions. Large incisor and canine tooth sockets indicate that the man probably had quite large front teeth and a broad mouth. The mandible of this individual is not known, but the phylogenetic analyses suggest that the Harbin cranium and the Xiahe mandible from Gansu Province of China form a sister group. The M2 size of the Harbin cranium matches the tooth size of the Xiahe mandible. It is reasonable to deduce that the Harbin cranium probably matches a mandible as robust as the Xiahe mandible and without a chin. It is hard to reconstruct the skin tone and hair color of the Harbin individual without genetic information, but available genetic data suggest that Neanderthals, Denisovans, and early H. sapiens generally had relatively dark skin, hair, and eye color. Considering the high latitude of the provenance of the Harbin cranium, we have chosen to give the reconstruction only a medium-dark skin color (Figure 3). |
|
|
|
Post by Admin on Jul 1, 2021 19:57:15 GMT
Phylogenetic position of the Harbin cranium Our extensive phylogenetic analyses based on parsimony criteria13 and Bayesian inference14, 15, 16, 17 firstly support the monophyly of Neanderthals and the monophyly of H. sapiens (Figures 4 and S19–S23). The Irhoud fossils from Morocco form the most basal operational taxonomic unit (OTU) of the H. sapiens clade, and the Sima de los Huesos crania from Spain form the most basal OTU of the Neanderthal clade, in line with other current interpretations.18, 19, 20 The Harbin cranium and Xiahe mandible form a sister group, and they, plus the Dali, Hualongdong, Jinniushan specimens, the European H. antecessor partial cranium, the African Eliye Springs cranium, and Rabat palate, form a monophyletic group. This clade forms the sister group of the similarly monophyletic H. sapiens clade. The specimens traditionally grouped in H. heidelbergensis/H. rhodesiensis do not constitute a monophyletic group and the Asian and African H. erectus specimens similarly form a paraphyletic group. When backbone constraints are used to reflect the results from palaeoproteomic and ancient DNA research by forcing the Xiahe mandible as the sister group of Neanderthals21 and H. antecessor outside of the H. sapiens-Neanderthal clade,22 Chinese late Middle Pleistocene humans, including the Harbin cranium, form a monophyletic clade as the sister group of Neanderthals (Figure S20). Both most parsimonious and backbone partially constrained phylogenetic trees support the monophyly of the group, including Dali, Jinniushan, Hualongdong, Xiahe, and Harbin.  Figure 4 Phylogeny of the 55 selected fossils from the genus Homo Some researchers have proposed that all Middle Pleistocene hominins belong to a single lineage leading to modern humans, with Asian Middle Pleistocene hominins, such as Dali and Hualongdong, suggested as transitional forms between Asian H. erectus and Asian H. sapiens specimens.6,9,24 Some other researchers have recognized these Asian hominins as part of the H. heidelbergensis/H. rhodesiensis hypodigm.25, 26, 27 A previous analysis based on overall similarity showed differences between Dali-Maba and the H. heidelbergensis hypodigm, and the potential connection between Dali-Maba and African early H. sapiens.28 Our analyses suggest that the Harbin cranium, together with Dali, Jinniushan, Hualongdong, and Xiahe, is not a part of the African and European H. heidelbergensis/H. rhodesiensis clade, but is the sister group of H. sapiens (see also the backbone partially constrained parsimony analysis in the supplemental information). The sister relationship between Harbin and Xiahe, as identified by Bayesian inference (but not parsimony analysis, see the supplemental information), is particularly interesting. The Xiahe mandible shows some proteomic features of the Denisovans,21 who were informally called “Homo sapiens altaiensis” or “Homo altaiensis,”12,29 and sediments from Baishiya Cave have yielded Denisovan mtDNA.30 The Harbin M2 also matches the known permanent Denisovan molars in size and root morphology, and, ever since the discovery of Denisovans, Asian Middle Pleistocene hominins, such as Dali, Jinniushan, and Xujiayao, have been suspected to represent an East Asian population of the Denisovans.31 More mandibular specimens for the Harbin population or cranial specimens corresponding to the Xiahe mandible will test how close the Harbin and Xiahe humans are morphologically, while new genetic material will test the relationship of these populations to each other and to the Denisovans. The results of the Bayesian tip-dating analyses suggest that the Harbin and Xiahe fossils shared a common ancestor ∼188 ka (397–155 ka), and the clade, including the Harbin cranium and H. sapiens shared a common ancestor at ∼949 ka (1,041.41–875.25 ka). The Neanderthal-H. sapiens divergence time in our analysis was ∼1,007 ka (1,114–919 ka). This estimation falls in the range based on mtDNAs for the split between the basal Neanderthal (Sima de los Huesos) and the H. sapiens lineage,20 but is much older than the estimation based on nuclear DNAs for the splits between the Neanderthal and H. sapiens lineages.32, 33, 34 However, it is possible that this younger estimated divergence date is an artifact of statistical averaging between “super-archaic” and “recent gene flow” events.35 The common ancestor of the H. sapiens OTUs included in our analysis is as old as ∼770 ka (922–622 ka), suggesting that the H. sapiens clade has a much deeper origin time than previously estimated. The Eurasian H. sapiens OTUs share a common ancestor ∼416 ka (534–305 ka) old. Outside of Africa, however, the earliest known H. sapiens fossil is only ∼210 ka.36 There is a large time gap between the hypothetical common ancestor of Eurasian H. sapiens and the actual fossil record, from the Bayesian tip-dating analysis. One plausible hypothesis is that the ancestral population of Eurasian H. sapiens may have diversified in Africa for many millennia before they dispersed into Eurasia. Genetic studies on ancient DNA suggest that the initial genetic exchanges between Neanderthals and H. sapiens occurred between 468 and 219 ka,33 or between ∼370 and 100 ka,34 and the introgression may have originated through gene flow from an African source.19,33 Interestingly, not only does the estimated time of the introgression event between Neanderthals and H. sapiens roughly overlap our prediction for the age of the common ancestor of Eurasian H. sapiens, but the African origin of the introgression is also consistent with our African ancestral population hypothesis. |
|
|
|
Post by Admin on Jul 1, 2021 22:13:54 GMT
Biogeography of the Homo species/populations We conducted maximum likelihood analysis under 18 different biogeographical models and estimated the number and type of biogeographical events using biogeographical stochastic mapping (BSM). The Akaike information criterion (AIC) model selection strongly supported dispersal-extinction cladogenesis37,38 with the founder-event dispersal (“jump dispersal”)39,40 model (DEC + j) as the best fit and the most probable biogeographical model (Tables S12–S14). Under this best fitting model (Figure 5), the ancestral distribution range of the Harbin, Dali, Jinniushan, Xiahe and Hualongdong group is most probably in Asia. The ancestral area for the Harbin-H. sapiens clade is most probably from Africa, supporting the idea that Africa is the center of origin of the H. sapiens clade. The ancestral distribution of the group bracketing Neanderthal, H. sapiens, and Harbin is from Africa or Europe.  Figure 5 Maximum likelihood ancestral range estimations and dispersal events for the Pleistocene Homo species/populations Our simulation of the biogeographical history of Homo species/populations identified sympatry diversification (∼57%) and founder-event dispersal (∼42%) as the common types of biogeographical modes across the phylogenetic tree of Homo (Table S15). Because all the OTUs are at the population level from a single locality, it is reasonable to find that no range expansion or range contraction event is detected from the BSM simulations. Founder-event dispersal usually involves a small number of individuals that dispersed to a new locality through a long dispersal distance and established a new isolated founder population.42, 43, 44 The changes in distribution range occurred at a lineage-splitting node, resulting in one daughter lineage dispersal into a new range, and the other daughter lineage remaining in the ancestral range. Sympatric diversification and founder-event dispersal being the most dominant biogeographical modes reflects the fact that multi-lineages of Homo coexisted in Africa, Europe, and Asia during the Middle and Late Pleistocene. These Homo lineages probably had a strong capability of dispersing for long distances, but remained in relatively small and isolated populations. BSMs indicate that the directionality of the dispersals between Africa, Asia, and Europe is asymmetric (Figure 5; Table S15). Asia is a sink of Homo species/populations that receives more dispersals from Africa and Europe than it gives dispersals to Africa and Europe. In total, Asia receives ∼42% of the total dispersal events and only provides ∼24% dispersals to other continents (Figure 5; Table S16). Africa is the major source of Homo dispersals. In total, ∼40% of all the dispersals are from Africa, while Africa also receives ∼22% dispersals from Asia and Europe. Instead of a unidirectional “out of Africa” model, a multi-directional “shuttle dispersal model” is more likely to explain the complex phylogenetic connections among African and Eurasian Homo species/populations. Conclusions The Harbin cranium is one of the best preserved of all archaic human fossils and its estimated late Middle Pleistocene age places it as an Asian contemporary of the evolving H. sapiens, H. neanderthalensis, and Denisovan lineages. It is huge in size, and its distinctive combination of traits in the cranial vault and face differentiate it from H. sapiens and H. neanderthalensis, as well as from the earlier species H. heidelbergensis/H. rhodesiensis. Instead it shows the greatest resemblances to Middle Pleistocene Chinese fossils, such as Hualongdong, Dali, and Jinniushan. This is confirmed by phylogenetic analyses using parsimony and Bayesian methods, which place these Chinese fossils with Harbin as a part of the sister group to H. sapiens, based on synapomorphies, such as a moderate post-toral sulcus, gently arched zygomaticoalveolar crest, presence of inferior orbital torus, strong malar tubercle, and thick mastoid processes. Our analyses also suggest a potential link between the Harbin cranium and the Xiahe mandible, a fossil attributed to the Denisovan lineage. The northerly location of the Harbin site also has implications for Middle Pleistocene human adaptive capabilities, since, even in the present interglacial, this region has winter temperatures averaging more than 16°C below zero. The very large size of the Harbin individual (as judged from the size of the cranium) may indicate physical adaptation to such conditions.45 The coexistence of several human lineages during the late Middle and Late Pleistocene of Asia is probably related to its diverse palaeoenvironments (ranging from the Gobi Desert to rainforest, and from coastal plains to the Qinghai-Tibet Plateau), which produced a varied biogeographic sink for human evolution. |
|
|
|
Post by Admin on Jul 2, 2021 2:47:39 GMT
‘Dragon Man’ may be an elusive Denisovan Ann Gibbons Science 02 Jul 2021: Vol. 373, Issue 6550, pp. 11-12 DOI: 10.1126/science.373.6550.11 Summary Almost 90 years after a Chinese bridge builder discovered a remarkably complete human skull and hid it in a well, Chinese scientists are now introducing it as "Dragon Man," the newest member of the human family, who lived more than 146,000 years ago. In three papers in the year-old journal The Innovation, paleontologist Qiang Ji of Hebei GEO University and his team describe the skull and argue it represents a new species that is a sister group to Homo sapiens, even closer kin to us than were the Neanderthals. Other researchers question that idea. But they suspect the large skull, which the team calls H. longi (long means dragon in Mandarin), has an equally exciting identity: They think it may be the long-sought skull of a Denisovan, an elusive human relative from Asia known chiefly from DNA. View Full Text: science.sciencemag.org/content/373/6550/11.full |
|
|
|
Post by Admin on Jul 5, 2021 20:15:43 GMT
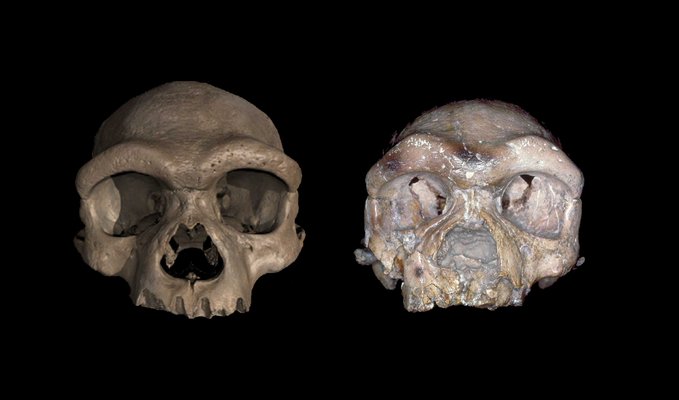 The reported discovery of a new hominin species from China created a lot of buzz last week. Its discoverers—paleoanthropologists Xijun Ni, Qiang Ji, Chris Stringer, and their colleagues—say that a skull discovered near Harbin, in northeast China, has a combination of features that's so different from Neanderthals, Denisovans, and Homo sapiens that it must be a separate species. The researchers have named the find Homo longi after the river where the skull was unearthed. Based on statistical comparisons of the skull's measurements with skulls from other hominins, Ni and colleagues say that Homo longi is a sister species to Neanderthals, Denisovans, and us. But that's still very much open for debate among paleoanthropologists, and the debate raises questions about how (or whether) we should draw lines between hominin species. Meet the Harbin skull Based on uranium-series dating, the Harbin skull lay buried for at least 146,000 years, but it's in remarkably good shape. Fossil hominin skulls often end up crushed or warped by the weight of the earth above them after many millennia in the ground, but the Harbin skull isn't distorted at all. It's also intact, even though the only tooth still attached is a left molar. That's unusual in itself, because teeth usually are the most common hominin fossil finds. The skull has a wide face with small, flat cheekbones, set underneath the dome of the skull. The face doesn't stick out in front of the skull like an australopithecine or a modern ape, although the upper jaw still protrudes a bit. Large eye sockets sit beneath heavy, curving brow ridges. The braincase is as large as a modern person's, but it's long and low rather than high and round. The jaw once held large molars that look like they belong to an older member of our family tree—or a Denisovan. Each of those individual features has been seen before in other hominin species. Species like Homo erectus and Homo heidelbergensis have long, low cranial vaults like the Harbin skull. But they also both have a bulge of bone called an occipital torus (or occipital bun) at the back of their skulls. Long, low skulls and occipital buns are both usually signs of an older branch of the human family tree: traits inherited from our last common ancestor with modern apes, rather than new ones evolved as our paths diverged. The Harbin skull's heavy brow ridges, protruding upper jaw, and big, robust molars are also considered archaic features. Old and new On the other hand, a relatively flat face that sits just beneath the brows, instead of sticking out front, is a hallmark of modern humans. So are the skull's small, low cheekbones and its large cranial capacity (about 1,420 milliliters, if you're counting). Then again, the Harbin skull is just impressively large in general. As Ni and colleagues wrote, "its enormous overall size sets it apart from nearly every other fossil," so we can't be sure how big its braincase was in relation to its body size. According to Ni and colleagues, anthropologists have never seen all of those features together in this exact combination until the Harbin skull. It's what paleoanthropologists call a mosaic, with some traits that look like they come from older lineages and some that seem more like ours. Mosaics are common in the hominin family tree. But the important question about the Harbin skull is whether its combination of features is really the trademark of a whole separate species, related to—but distinct from—Neanderthals, Denisovans, and us. Paleoanthropologists have unearthed several Neanderthal skulls, but the only Denisovan fossils known so far are a few teeth, a finger bone, and a single skull fragment from Denisova Cave in Siberia as well as a jawbone from Xiahe in northern China. In other words, we don't have many examples to compare the Harbin mandible to if we want to say whether it looks like a Denisovan skull. There's no shortage of Homo sapiens skulls to look at, of course, and it's clear that, whatever the Harbin skull once was, it didn't belong to a member of our species (spoiler alert: depending on how we define our species). Looking at skulls from modern humans, however, should remind us how much diversity in size, shape, and appearance can exist within a species, especially one that's adapted to live in environments all over the world. The Harbin cranium could be well within the normal range of diversity for Denisovans, who were also a very widespread species, or it could be something completely different. We just don't have enough information about Denisovan skulls to know yet. Denisovan or Homo longi? When Ni and colleagues did their statistical analysis, they pointed out that the Harbin skull fell into a group along with the 160,000-year-old Denisovan mandible from Xiahe, which anthropologists identified based on proteins still preserved within it. In other words, Harbin and Xiahe probably belong to the same species or to closely related species. Meanwhile, the one molar still attached to the Harbin skull looks remarkably like two Denisovan molars from Denisova Cave in Siberia. The Harbin skull also fits in well with a 200,000-to-260,000-year-old skull found in Dali County in northwestern China, a roughly 300,000-year-old skull found in Hualong Cave in eastern China, and a 260,000-year-old skull from Jinniushi (sometimes spelled Jinniushan) Cave in China. All three of those fossils have been described as looking like something in between Homo erectus and modern humans. It's possible—although still mostly fodder for speculation—that all or some these fossils, along with some from Taiwan and northern China whose molars look an awful lot like the Xiahe ones, could turn out to be an East Asian group of Denisovans. That would still make the Harbin cranium an important find. It doesn't have to represent a brand-new-to-science species to be a valuable piece of a mostly unwritten chapter of our evolutionary story. Denisovan remains are still extremely scarce, and it's increasingly clear that East Asia was an important place for our ancestors during the Early and Middle Paleolithic. Then again, at least some of the Asian hominin fossils might also turn out to be, as Ni and colleagues claim, a distinct species. Only more evidence can clear up that debate, and Ni and colleagues acknowledge that in one of their recent papers. "More mandibular specimens for the Harbin population, or cranial specimens corresponding to the Xiahe mandible, will test how close the Harbin and Xiahe humans are morphologically," they wrote, "while new genetic material will test the relationship of these populations to each other and to the Denisovans." |
|

