|
|
Post by Admin on Dec 24, 2021 19:41:36 GMT
Y-chromosome haplogroups from Hun, Avar and conquering Hungarian period nomadic people of the Carpathian Basin Abstract Hun, Avar and conquering Hungarian nomadic groups arrived to the Carpathian Basin from the Eurasian Steppes and significantly influenced its political and ethnical landscape, however their origin remains largely unknown. In order to shed light on the genetic affinity of above groups we have determined Y chromosomal haplogroups and autosomal loci, suitable to predict biogeographic ancestry, from 49 individuals, supposed to represent the power/military elit. Haplogroups from the Hun-age are consistent with Xiongnu ancestry of European Huns. Most of the Avar-age individuals carry east Eurasian Y haplogroups typical for modern north-eastern Siberian and Buryat populations and their autosomal loci indicate mostly un-admixed Asian characteristics. In contrast the conquering Hungarians seem to be a recently assembled population incorporating un-admixed European, Asian as well as admixed components. Their heterogeneous paternal and maternal lineages indicate similar supposed phylogeographic origin of males and females, derived from Central-Inner Asian and European Pontic Steppe sources.  The Bashkir R1b-M73 (R1b1a1a1) descend directly from the Bronze-age Proto-Indo-Europeans. The only place where both M73 and M269 are both common is around the Caucasus and Anatolia. In the Pontic steppes during the Neolithic, the first steppe invaders would have belonged to both R1b-M73 (R1b1a1a1) and R1b-M269 (R1b1a1a2; previously R1b1a2), although the latter would have been much more dominant. It is possible that all the subclades as far as S116 and even S28/U152 developed in the steppes before migrating to Europe. The Bashkirs could represent the last leftovers from these PIE R1b subclades. Lobov et al. (http://ibg.anrb.ru/Lobov_AS.PDF) has the haplogroup distribution for the Bashkirs in 8 locations. R1b-M73 ranges from 0% to 55%. R1b-M269 ranges from 0% to 84%.  Break down for R1b subclades in Bashkirs n* M73 M269(xL23) L23(xM412) U152 South-East Baskkirs 329 77 8 106 2 West Bashkirs 54 0 0 0 0 South Bashkirs 79 0 2 9 1 North Bashkirs 70 1 0 2 50 Southe-West Baskkirs 51 1 0 0 0 Total 586 79 10 126 53 * number of sample Bashkirs |
|
|
|
Post by Admin on Dec 24, 2021 20:38:58 GMT
Introduction Population history of the Carpathian Basin was profoundly determined by the invasion of various nomadic groups from the Eurasian Steppes during the Middle Ages. Between 400–453 AD the Huns held possession of the region and brought about a major population reshuffling all over Europe. Recent genetic data connect European Huns to Inner Asian Xiongnus1, but genetic data from Huns of the Carpathian Basin have not been available yet, since Huns left just sporadic lonely graves in the region, as they stayed for short period. We report three Y haplogroups (Hg) from Hun age remains, which possibly belonged to Huns based on their archaeological and anthropological evaluation.  From 568 AD the Avars established an empire in the region lasting nearly for 250 years, until they were defeated by the Franks and Bulgars in 803, then their steppe-empire ended around 822 AD. In its early stage the Avar Khaganate controlled a large territory expanding from the Carpathian Basin to the Pontic-Caspian Steppes and dominated numerous folks including Onogur-Bulgars, which fought their independence in the middle 7th century and established the independent Magna Bulgaria state. Then the Avar Khaganate was shrunken, its range well corresponding to that of the succeeding Hungarian Kingdom. The Avars arrived in multiple waves into the Carpathian Basin and the Avar period left a vast archeological legacy with more than 80 thousand excavated graves in present-day Hungary. The Avar age remains are anthropologically extremely heterogeneous, with considerable proportion of Mongoloid and Europo-Monoloid elements reaching 20–30% on the Great Hungarian Plain2, attesting that the Carpathian Basin witnessed the largest invasion of people from Asia during this period. Most individuals buried with rich grave goods show Mongoloid characters indicating inner Asian origin of the Avar elite, which is also supported by their artifact types, titles (e.g. khagan) and institutions recognized to be derived from Inner Asian Rourans. From the Avar period only a few mitochondrial DNA (mtDNA) data are available from two micro-regions3,4, which showed 15.3% and 6.52% frequency of East Eurasian elements. A recent manuscript described 23 mitogenomes from the 7th-8th century Avar elite group5 and found that 64% of the lineages belong to East Asian haplogroups (C, D, F, M, R, Y and Z) with affinities to ancient and modern Inner Asian populations corroborating their Rouran origin. Though the Avar Khaganate ceased to exist around 822 AD, but its population survived and were incorporated into the succeeding Hungarian state6. It is relevant to note that none of the Hungarian medieval sources know about Avars7, probably because they were not distinguished from the Huns as many foreign medieval sources also identified Avars with Huns, for example the Avars who were Christianized and became tax-payer vassals of the Eastern Frankish Empire were called as Huns in 8718. Presence of the Hungarians in the Carpathian Basin was documented from 862 AD and between 895–905 they took full command of the region. The Hungarians formed a tribal union but arrived in the frame of a strong centralized steppe-empire under the leadership of prince Álmos and his son Árpád, who were known to be direct descendants of the great Hun leader Attila, and became founders of the Hungarian ruling dynasty and the Hungarian state. The Hungarian Great Principality existed in Central Europe from ca. 862 until 1000, then it was re-organized as a Christian Kingdom by King István I the Saint who was the 5th descendant of Álmos9. Our recent analysis of conquering Hungarian (hence shortened as Conqueror) mitogenomes revealed that the origin of their maternal lineages can be traced back to distant parts of the Eurasian steppe10. One third of the maternal lineages were derived from Central-Inner Asia and their most probable ultimate sources were the Asian Scythians and Asian Huns, while the majority of the lineages most likely originated from the Bronze Age Potapovka-Poltavka-Srubnaya cultures of the Pontic-Caspian steppe. Population genetic analysis indicated that Conquerors had closest connection to the Onogur-Bulgar ancestors of Volga Tatars. Although mtDNA data can be informative, since as far as we know none of the above nomadic invasions were pure military expeditions in which raiding males took local women, but entire societies with both men and women migrated together3, however nomadic societies were patrilinealy organized, thus Y-chromosome data are expected to provide more relevant information about their structure and origin than mtDNA. So far 6 Y-chromosome Hg-s have been published from the Conquerors11; which revealed the presence of N1a1- M46 (previously called Tat or N1c), in two out of 4 men, while12 detected two R1b-U106 and two I2a-M170 Hg-s. In order to generate sufficient data for statistical evaluation and compare paternal and maternal lineages of the same population, we have determined Y-Hg-s from the same cemeteries whose mtDNA Hg-s were described in10. As most Hungarian medieval chronicles describe the conquer as the “second incoming of the Hungarians”7, genetic relations may exist between consecutive nomadic groups, first of all between elite groups. As the studied Conqueror cemeteries represent mainly the Conqueror elite, we supplemented our Y-Hg studies with samples from the preceding Avar power elite/millitary elite groups and available samples from the Hun period to test possible genetic relations. Avar military leader graves are distinguished by high-value prestige artifacts: belt decorated with gold or silver mounts, decorated horse harness, saddle, sword, bow and quiver with arrowheads, gold decorated plates, silver or gold earrings (Supplementary Information). Conqueror leader graves contain characteristic partial horse burial with horse cranium and leg bones or symbolic horse burial with adorned harness, saddle and stirrup. Elite grave goods often include precious metal jewels, belt with impressed metal belt-buckles, metal-plated sabretache and weaponry; arrowheads, quiver and bow10,12. From the studied samples we also tested autosomal single nucleotide polymorphisms (SNP-s) associated with eye/hair/skin color and lactase persistence phenotypes as well as biogeographic ancestry. |
|
|
|
Post by Admin on Dec 24, 2021 21:40:36 GMT
Results We selected 168 phylogenetically informative Y chromosome SNP-s13 defining all major Hg-s and the most frequent Eurasian sub-Hg-s, as well as the following 61autosomal SNP-s: 25 HirisPlex markers suitable for eye and hair color prediction14, two SNP-s linked to adult lactase persistence15 and 34 ancestry informative markers (AIMs)16, as listed in Supplementary Tables S1 and S2. DNA segments containing above SNP-s were enriched from the next generation sequencing (NGS) library of each sample with hybridization capture. Anthropological males were selected from the cemeteries, but as control we also included four females for which the presence of Y-chromosomal reads can reveal contamination. Analysis of women control samples revealed low contamination, having negligible Y-chromosomal reads but comparable autosomal reads to males (Supplementary Tables S1 and S2) with one exception, KEF1/10936 genetically turned out to be a male despite its anthropological description as female (Supplementary Table S6). Negligible contamination levels for 26 of the 49 libraries had been demonstrated before10 and low contamination is also inferred from unambiguous Hg classifications with very few contradicting SNP-s, most of which can be explained by postmortem ancient DNA modifications (Supplementary Table S1). Besides, transition patterns of DNA molecules and library fragment size distributions were typical for ancient DNA and we also estimated low contamination by calculating the proportion of reads which did not correspond to the consensus sequence in diagnostic positions (Supplementary Table S3). Moreover we detected a significant number of east Eurasian Hg-s which had a negligible chance to have been derived from a contaminating source of European researchers. We obtained low to medium average coverage of the targeted loci, (Supplementary Tables S1, S2 and S3) and Y-Hg determination was consistent from 46 males (Supplementary Table S1) which is summarized in Fig. 1 and phylogeographic distribution of Conqueror lineages is displayed on Fig. 2. Figure 1 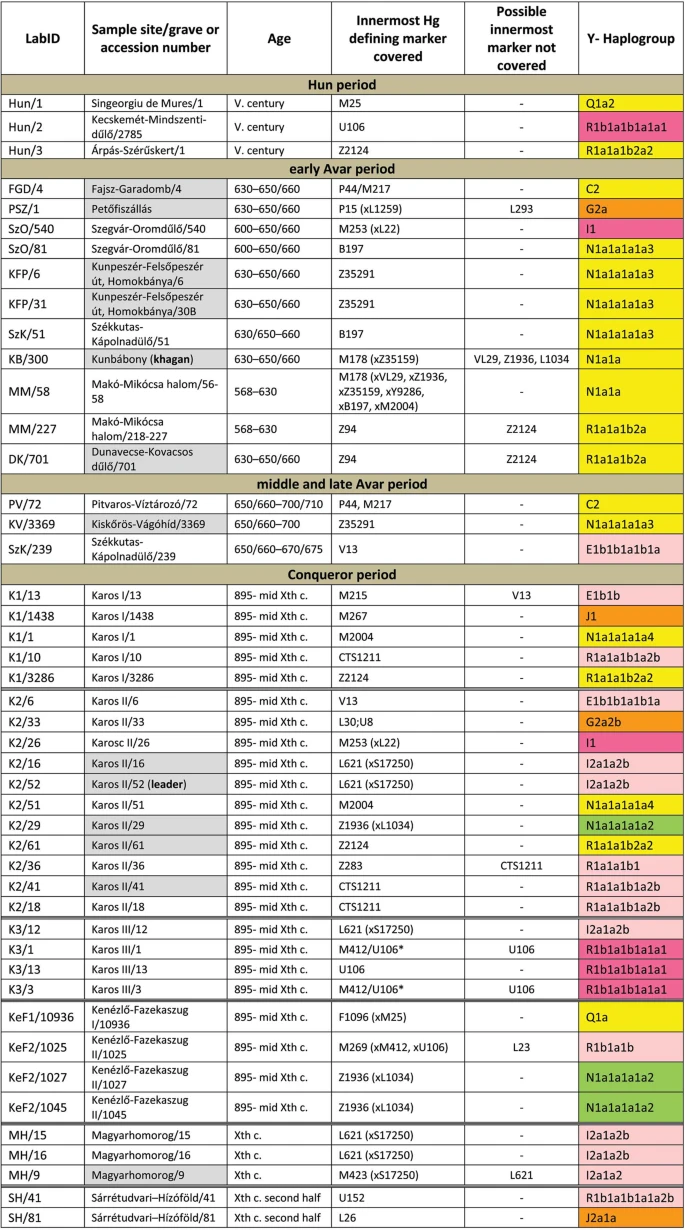 Y-Hg-s determined from 46 males grouped according to sample age, cemetery and Hg. Hg designations are given according to ISOGG Tree 2019. Grey shading designate distinguished individuals with rich grave goods, color shadings denote supposed geographic origin of Hg-s according to Fig. 2. For samples K3/1 and K3/3 the innermost Hg defining marker U106* was not covered, but had been determined previously12. Figure 2 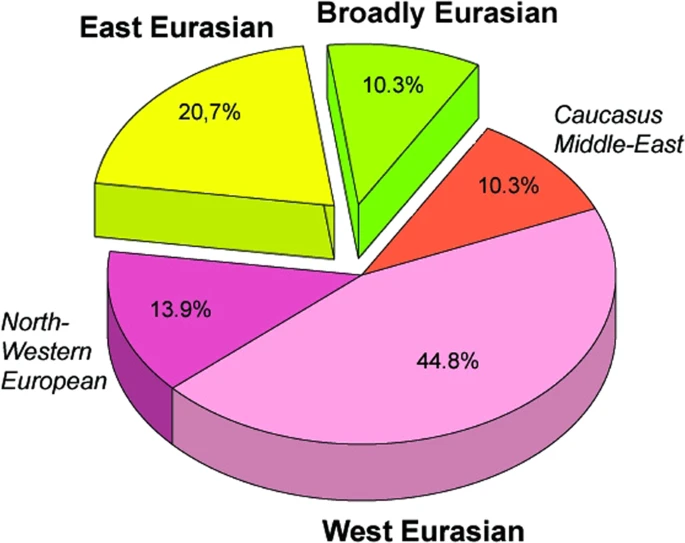 East Eurasian Hg-s Hg Q1a2- M25 is very rare in Europe, where it has highest frequency among Seklers (a Hungarian speaking ethnic group in Transylvania) according to Family Tree DNA database. Ancient samples with Hg Q1a2- M25 are known from the Bronze Age Okunevo and Karasuk cultures, as well as Middle Age Tian Shan Huns and Hunnic-Sarmatians17 implying possible Hunnic origin of this lineage in Europe, which is confirmed by the Hg of our Hun/1 sample, derived from Transylvania. One Conqueror sample KEF1/10936 belongs to Q1a- F1096, possibly to the Q1a1-F746 sister clade of Q1a2- M25, which was not tested in our experiment. Q1a1 is present in East Asia; Mongolia, Japan, China, Korea and its presence in the Kenézlő (KEF) graveyard imply a common substrate of Huns and Conquerors. Hun/3 belongs to Hg R1a1a1b2a2- Z2124, a subclade of R1a1a1b2-Z93, the east Eurasian subbranch of R1a. Today Z2124 is most frequent in Kyrgyzstan and Afghanistan, but is also widespread among Karachai-Balkars and Baskhirs18. Z2124 was widespread on the Bronze Age steppe, especially in the Afanasievo and Sintashta cultures19 and R1a detected in Xiongnus20,21 very likely belong to the same branch. Two samples from the Karos Conqueror cemeteries (K1/3286 and K2/61) were also classified as R1a-Z2124 and two Avar age individuals (DK/701 and MM/227) belong to the same R1a1a1b2a-Z94 branch but marker Z2124 was not covered in latter samples. Two Avar samples belonged to Hg C2-M217, which is found mostly in Central Asia and Eastern Siberia. Presence of this Hg had also been detected in the Conquerors, as the Karos2/60 individual belongs to C2 (Fóthi unpublished). All N-Hg-s identified in the Avars and Conquerors belonged to N1a1a-M178. We have tested 7 subclades of M178; N1a1a2-B187, N1a1a1a2-B211, N1a1a1a1a3-B197, N1a1a1a1a4-M2118, N1a1a1a1a1a-VL29, N1a1a1a1a2-Z1936 and the N1a1a1a1a2a1c1-L1034 subbranch of Z1936. The European subclades VL29 and Z1936 could be excluded in most cases, while the rest of the suclades are prevalent in Siberia22 from where this Hg dispersed in a counter-clockwise migratory route to Europe23. Avar sample MM/58, did not go into any of the tested M178 subclades, while only N1a1a2 could be excluded for the KB/300 Avar khagan due to low coverage. All the 5 other Avar samples belonged to N1a1a1a1a3-B197, which is most prevalent in Chukchi, Buryats, Eskimos, Koryaks and appears among Tuvans and Mongols with lower frequency22. By contrast two Conquerors belonged to N1a1a1a1a4-M2118, the Y lineage of nearly all Yakut males, being also frequent in Evenks, Evens and occurring with lower frequency among Khantys, Mansis and Kazakhs. Broadly Eurasian Hg-s Three Conqueror samples belonged to Hg N1a1a1a1a2-Z1936, the Finno-Permic N1a branch, being most frequent among northeastern European Saami, Finns, Karelians, as well as Komis, Volga Tatars and Bashkirs of the Volga-Ural region. Nevertheless this Hg is also present with lower frequency among Karanogays, Siberian Nenets, Khantys, Mansis, Dolgans, Nganasans, and Siberian Tatars22. West Eurasian Hg-s
The west Eurasian R1a1a1b1a2b-CTS1211 subclade of R1a is most frequent in Eastern Europe especially among Slavic people. This Hg was detected just in the Conqueror group (K2/18, K2/41 and K1/10). Though CTS1211 was not covered in K2/36 but it may also belong to this sub-branch of Z283.Hg I2a1a2b-L621 was present in 5 Conqueror samples, and a 6th sample form Magyarhomorog (MH/9) most likely also belongs here, as MH/9 is a likely kin of MH/16 (see below). This Hg of European origin is most prominent in the Balkans and Eastern Europe, especially among Slavic speaking groups. It might have been a major lineage of the Cucuteni-Trypillian culture and it was present in the Baden culture of the Calcholitic Carpathian Basin24. I1- M253, identified from one Conqueror sample is a northern European Hg found mostly in Scandinavia and Finland and might have originated from this region during the Mesolithic. It has somewhat similar distribution to R1b-U106 associated with Germanic speaking populations. Three out of 4 samples in the small Karos3 cemetery belonged to Hg R1b1a1b1a1a1-U106 setting apart this cemetery from all other groups, except for the Hun/2 sample which is the only other one with this Hg. Hg U106 is considered a “Germanic” branch as it is most significant today in Germany, Scandinavia, and Britain, and rare in Eastern Europe (Supplementary Table S4). Its ancestral branch Hg R1b1a1b-M262 is assumed to have emerged in the Pontic-Caspian Steppe and arrived to Europe with Bronze Age migrations25. Its presence in Hun and Conqueror samples may derive from Goths, Gepids or other German allies of the Huns.
We detected R1b1a1b1a1a2b-U152 in one sample from the Sárrétudvari Conqueror cemetery, which represent rather commoners than the elite. U152 is the Italo-Celtic R1b branch, concentrated around the Alps, and which was present in the Carpathian Basin before the conquer, so did not necessarily arrived with the Conquerors.The mediterranean haplogroup E1b1b1a1b1a-V13 was detected in an Avar (SzK/239) and a Conqueror (K2/6) sample, while this marker was not covered in another sample (K1/13, E1b1b- M215). This Hg originated in the Middle East and migrated to the Balkans and Western Asia during the Bronze Age. The PSZ/1 Avar leader belongs to Hg G2a-P15, while K2/33 to the G2a2b-L30 subbranch. Hg G2a is originated from Anatolia/Iran, now it is most common in the Caucasus region and its arrival to Europe is associated with the spread of Neolithic farmers26. One Conqueror sample belongs to Hg J1-M267 and another to J2a1a-L26. Both J1 and J2 lineages are most frequent around the Middle East-Caucasus and probably originated from this region27, then expanded with pastorists prior to the Neolithic. |
|
|
|
Post by Admin on Dec 25, 2021 0:10:56 GMT
Autosomal SNP-s We have predicted eye, hair and skin color phenotypes from 25 HirisPlex SNP-s, also suitable to predict non-European ancestry14, as summarized in Figs 3 and 4. Samples from different archaeological cultures and cemeteries showed a remarkable pattern of phenotypic distribution. All Hun and Avar age samples had inherently dark eye/hair colors, DK/701 being the only exception (Fig. 3). Moreover 6/14 Avar age samples were characterized with >0,7 black hair; >0,99 brown eye p-values, inferring 86,5% probability of non-European biogeographic ancestry14 in agreement with their anthropological, archaeological and historical evaluation. In contrast the Conquerors showed a wide variety of phenotypes clustered by cemeteries (Fig. 4). All individuals from the Sárrétudvari (SH), Magyarhomorog (MH) and majority from the Kenézlő (KEF) graveyards displayed European phenotypic patterns; blue eye and/or light hair with pale skin. In the three Karos cemeteries darker eye/hair colors predominated, 4/20 individuals having p-values consistent with non-European origin, nevertheless 5/20 individuals had light hair color indicating a rather mixed origin of this population, concurrent with their mtDNA and Y chromosomal Hg composition. 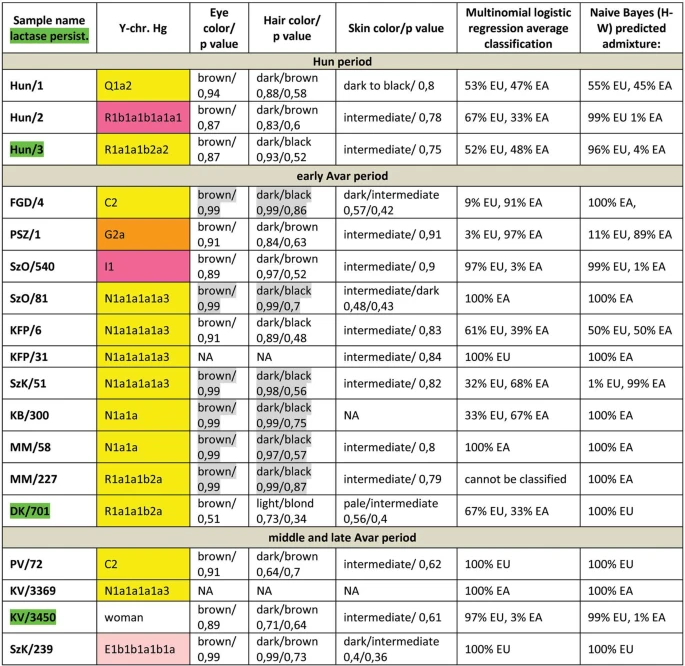 Figure 3 Phenotypes and predicted genetic origin of Hun and Avar age individuals. Eye, hair and skin colors with their probability values were predicted from HIrisPlex markers (Supplementary Table S2), gray shaded values predict non-European ancestry. Most likely origin of individuals was calculated from 34 AIMs (Supplementary Table S2) with two methods; multinomial logistic regression and naive Bayesian classifier (supposing Hardy-Weinberg equilibrium). “Cannot be classified” indicates that maximum probabilities of most likely ancestral origin are below 0.34. Names of individuals carrying SNP variants associated with lactase persistence are highlighted with green. Abbreviations are the following: NA = lack of data, EU = Europe, EA = East Asia AF = Africa. 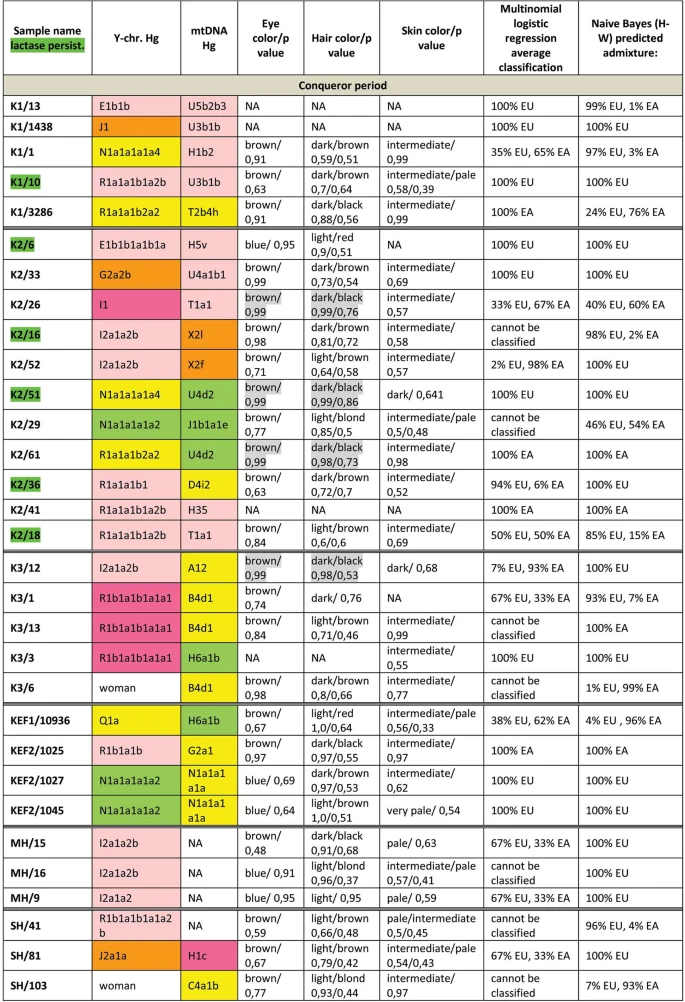 Figure 4 Phenotypes and genetic origin of Conqueror period individuals. mtDNA Hg-s were taken from10, all markings are identical to that of We have also determined 34 AIMs, which can inform about the likely geographic origin of individuals, and used the Snipper App suite version 2.5 portal16 to assign biogeographic ancestry. We predicted ancestry with both the naive Bayesian classifier and multinomial logistic regression (MLR) algorithms, as these make different assumptions about genetic equilibrium28, and listed the results on Figs 3 and 4. The AIM-s results fairly matched and complemented phenotypic information. All Hun age individuals revealed admixture derived from European and East Asian ancestors, while 8/15 Avar age individuals showed predominantly East Asian origin with both methods, 4 individuals were definitely European, while two showed evidence of admixture. The KFP/31 sample gave contradicting results due to low coverage. Conqueror samples from the Magyarhomorog (MH) and Sárrétudvari (SH) cemeteries showed mostly European ancestry in agreement with their phenotypes and Y Hg-s, though MLR detected a significant east Asian ancestry component and the SH/103 women was classified east Asian despite her blond hair. The Karos (K) and Kenézlő (KEF) populations were profoundly admixed, comprising individuals of purely East Asian, European and mixed origin in nearly identical proportions, again in agreement with results obtained from uniparental and phenotypic markers. The determined variable autosomal loci are also suitable to exclude possible direct (parent-child, sibling) genetic relatedness29, thus we compared the autosomal genotypes of all individuals sharing either maternal or paternal Hg-s. Direct kinship could not be excluded between the MH/9 and MH/16 individuals with identical phenotypes, suggesting that MH/9 probably also belongs to Hg I2a1a2b-L621, despite its uncovered L621 marker. KEF2per1027 and KEF2per1045 were probably brothers as they had identical mitogenomes, Y Hg-s and blue eye color besides sharing autosomal alleles. The same applies to K3/1 and K3/13 individuals who were probably also brothers. We could not exclude possible direct paternal relationship between K2/36, K2/18 and K2/41 but the first two samples had unsatisfactory coverage to make a strong statement. We have tested two SNP-s (rs4988235 and rs182549) associated with the adult lactase persistence phenotype in Europe15. Individuals carrying derived alleles in these loci are able to digest lactose in dairy products during adulthood without symptoms of lactose intolerance. Allele frequency of the persistence genotype varies throughout Eurasia30, reaching above 90% at some parts of Northwestern Europe, around 80% in present day Hungary, but drops below 30% in Central Asia and even lower in East Asia. We detected the derived persistence allele in all of the studied groups (Figs 3 and 4, Supplementary Table S2); 1/3 of the Hun period individuals, 2/14 of the Avar period individuals, and 6/31 of the Conqueror period individuals carried persistence alleles. It is remarkable that the persistence genotype seems to be associated with European origin, as all of the carriers were predicted to have predominantly European ancestors. This is in agreement with a previous study31, which found that all of the 11% Conqueror samples with persistence genotype carried European mtDNA Hg H. In addition all carriers were heterozygous, 6 of them for both SNP-s, but three of them carried just the rs182549 (-22018 G > A) derived allele suggesting previous admixture with non-carriers, possibly derived from East Eurasia.
|
|
|
|
Post by Admin on Dec 25, 2021 3:13:24 GMT
Population genetic analysis The studied Conqueror group very likely represent real populations, as 24 of the 29 samples came from 4 nearby cemeteries (Karos 1,2,3 and Kenézlő) with identical archaeological and anthropological groupings and the studied Magyarhomorog individuals were also categorized as belonging to the same early Conqueror elite. The Avar group was assembled from several different cemeteries of a wider timespan, thus they cannot represent the Avar period population of the Carpathian Basin, however their relatively homogenous Hg distribution indicate that the Avar elite embodied the same east Eurasian sub-population throughout their reign, so it appeared to be meaningful to include them in the analysis. In order to find the most similar populations to our studied ones, we compared the Hg distribution of the Avar and Conqueror elite to that of 78 modern Eurasian populations (Supplementary Table S4) and represented their relations on MDS plot displayed on Fig. 5. Only one individual was included in the analysis from first degree relatives. In order to increase resolution, we separately calculated the frequencies of each sub-Hg-s which are present in our samples, while frequencies of all other subclades were combined as listed in Supplementary Table S4. Similar Hg distributions are mapped into neighboring positions on the MDS plot, which clearly separates populations according to their geographic locations. Along the x axis east Eurasian populations map to the right while Europeans are compressed at the left. Along the y axis Siberian populations are sequestered at the top, while East Asian ones at the bottom of the graph. The Avars are obviously mapped to the Siberian side with smallest weighted Euclidean distances from Koryaks (KRY), Teleuts (TEL), Khantys (KHT), Komis, (KOM) and Dolgans (DLG). The Conquerors are positioned between eastern Europeans, Central Asians and Siberians but their exact relations are hard to make out because of the crowding at the European side. Figure 5 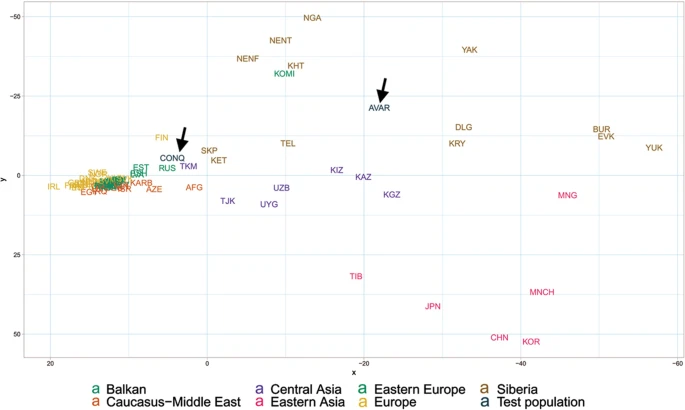 In order to better discern the position of the Conquerors we redraw the MDS plot without the eastern Asian and Siberian populations (Fig. 6). Though having removed subset of the data MDS imaging in two-dimensions inevitably rearranged the remaining components, but the general organization of the population clusters remained unchanged: Western-Northern Europeans map to the left, Central-eastern European and Balkan people around the middle, Central Asians to the top right, and Caucasus-Middle East populations to bottom right. Conquerors remained between Central Asians and eastern Europeans, with smallest weighted Euclidean distances from Bashkirs (BSH), Hungarians (HUN), Tajiks (TJK), Estonians (EST), Kazakhs (KAZ), Uzbeks (UZB) and Slovaks (SVK). 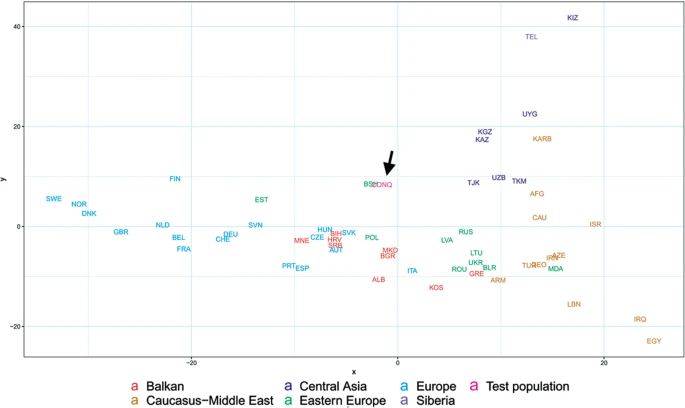 Figure 6 MDS plot of Y-chromosomal Hg distribution of 58 European and Central Asian populations including Conquerors (arrow). Population three letter codes are given in Supplementary Table S4. |
|

