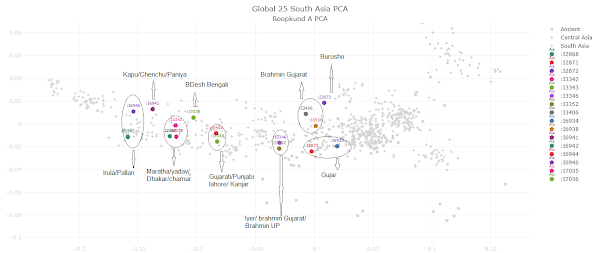
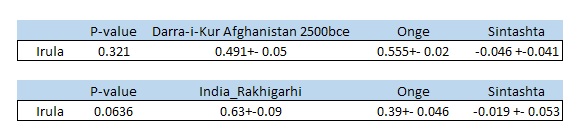
Post by Admin on Dec 30, 2021 21:50:36 GMT
Regarding the mtDNA analyses, our findings indicate
that the ancient Krasnoyarsk mtDNA pool harbored both
western and eastern Eurasian lineages. Nevertheless, most
of the retrieved sequences (n = 20, 77%) belong to western
Eurasian mtDNA haplogroups (HV, H, T, I, U and K). The
eastern Eurasian lineages (23% of the sequences) were represented
by haplogroups or subhapologroups C, Z, G2a,
F1b and N9a. The western Eurasian contribution to the
ancient mtDNA pool reached 90% for the Bronze Age and
decreased to 67% for the Iron Age. Thus, despite a small
sample size, our data suggests a temporal pattern which is
in agreement with the view that west Eurasian populations
predominated in the Krasnoyarsk region during the Bronze
Age, whereas Asian component began to increase from the
Iron Age on. This result is similar to that obtained in the
ancient DNA study of Lalueza-Fox et al. (2004) who
showed that all Kazakh sample specimens from before thirteenth
to seventh centuries BC belonged to European lineages. After
that time, there was an influx of East Asian
sequences which are thought to have coexisted with the
prior west Eurasian genetic substratum.
As shown in Table 5, and particularly in Fig. 3, the current
distribution of the ancient mtDNA haplotypes can be
broadly divided into three different geographic poles. The
first is represented roughly by eastern and northern Europe,
the second by the Volga–Ural region and the third by southern
Siberia. It is interesting to note that the distribution of
the paternal and maternal lineages is close. Indeed, except
for the Volga–Ural region, both maps overlap. This would
mean that the story of women matches well that of men. In
other words, the migrations in which south Siberian specimens
were involved seemed to be “whole-population
movements” rather than “war-like movements” involving
the men only. The fact that East Asian mtDNA sequences
appeared at the Iron Age could signify that once settled,
migrants of supposed European ancestry began to establish
relationships with groups coming from the east and to take
Asian women as wives. Moreover, the relative high diversity of the
mtDNA gene pool observed in the ancient specimens indicates that
numerous populations carrying different
mtDNA variants were involved in the formation of southern Siberian
populations, even reflecting long-distant
movements. It would not have presented any major difficulty
for Bronze Age and Early Iron Age peoples to range
from one end of Eurasia to the other within some centuries.
Historical records and archaeology attest that nomadic
groups moved across Eurasia from North of the Black sea,
through Central and Inner Asia, to northeast Asia in a matter of
centuries (Mair 2005). Some of them are described in
Chinese historiography as horse-riding, Caucasian-looking,
Indo-European-speaking people and are sometimes referred
as the “Kurgan Culture” (Zerjal et al. 2002). Paleogeographic
studies provide material which suggests that climate change,
particularly in the eastern regions of the
steppes, was among the causes of these population movements (Van Geel et al. 2004).
If we consider that there is a correspondence between the
overall distribution of haplotypes and haplogroups and past
human movements, it seems that the European or Caucasoid component
observed in the ancient Siberian sample
may originate from East European populations. Moreover,
it is likely that some mtDNA lineages were carried to
southern Siberia from the Volga–Ural region. Incidentally,
in the Wfth century BC, Herodotus mentioned transit trade
occurring in Central Asia along a route that stretched from
the Urals in the west to the Altai and the Minusinsk Basin
in the east (Hemphill and Mallory 2004). In Altai, the presence
of the R1a1 haplogroup in the middle of the fifth century BC is
confirmed by the sample SEB 96K2 of Ricaut
et al. (2004) which was found to belong to this Y-haplogroup.
The boundary of the eastern European influence
seems to be fixed at the peri-Baikal area since no R1a1
haplogroup was found in the Xiongnu specimens of the
Northern border of Mongolia.
According to the “Kurgan hypothesis” of Marija Gimbutas,
nomadic peoples of the Volga steppe region, assumed
to speak a Proto-Indo-European language, infiltrated
Europe in three waves between 4400 and 2800 BC. Around
4400 BC, Kurgan people from the lower Dnieper and lower
Volga regions began moving along the Black Sea littoral
into the Danube Basin. They migrated in the Central Balkans
and further into Central Europe. During the middle of
the fourth millennium BC, the Kurgan culture in the North
Pontic Region continued to develop. People travelled
across western Ukraine north of the Carpathian Mountains
to Poland and Central Germany. They also moved southwest into
eastern Romania. Shortly after 3000 BC, the third
Kurgan wave (Yamna people), originating once more from
the Volga steppe, spread from Central Europe to Northwest
Germany, the east Baltic area, southern Scandinavia, the
upper Dnieper basin and Central Russia. These three waves
of migrations might explain the distribution of mtDNA and
Y-chromosome lineages observed in the present work
(Figs. 2, 3)
that the ancient Krasnoyarsk mtDNA pool harbored both
western and eastern Eurasian lineages. Nevertheless, most
of the retrieved sequences (n = 20, 77%) belong to western
Eurasian mtDNA haplogroups (HV, H, T, I, U and K). The
eastern Eurasian lineages (23% of the sequences) were represented
by haplogroups or subhapologroups C, Z, G2a,
F1b and N9a. The western Eurasian contribution to the
ancient mtDNA pool reached 90% for the Bronze Age and
decreased to 67% for the Iron Age. Thus, despite a small
sample size, our data suggests a temporal pattern which is
in agreement with the view that west Eurasian populations
predominated in the Krasnoyarsk region during the Bronze
Age, whereas Asian component began to increase from the
Iron Age on. This result is similar to that obtained in the
ancient DNA study of Lalueza-Fox et al. (2004) who
showed that all Kazakh sample specimens from before thirteenth
to seventh centuries BC belonged to European lineages. After
that time, there was an influx of East Asian
sequences which are thought to have coexisted with the
prior west Eurasian genetic substratum.
As shown in Table 5, and particularly in Fig. 3, the current
distribution of the ancient mtDNA haplotypes can be
broadly divided into three different geographic poles. The
first is represented roughly by eastern and northern Europe,
the second by the Volga–Ural region and the third by southern
Siberia. It is interesting to note that the distribution of
the paternal and maternal lineages is close. Indeed, except
for the Volga–Ural region, both maps overlap. This would
mean that the story of women matches well that of men. In
other words, the migrations in which south Siberian specimens
were involved seemed to be “whole-population
movements” rather than “war-like movements” involving
the men only. The fact that East Asian mtDNA sequences
appeared at the Iron Age could signify that once settled,
migrants of supposed European ancestry began to establish
relationships with groups coming from the east and to take
Asian women as wives. Moreover, the relative high diversity of the
mtDNA gene pool observed in the ancient specimens indicates that
numerous populations carrying different
mtDNA variants were involved in the formation of southern Siberian
populations, even reflecting long-distant
movements. It would not have presented any major difficulty
for Bronze Age and Early Iron Age peoples to range
from one end of Eurasia to the other within some centuries.
Historical records and archaeology attest that nomadic
groups moved across Eurasia from North of the Black sea,
through Central and Inner Asia, to northeast Asia in a matter of
centuries (Mair 2005). Some of them are described in
Chinese historiography as horse-riding, Caucasian-looking,
Indo-European-speaking people and are sometimes referred
as the “Kurgan Culture” (Zerjal et al. 2002). Paleogeographic
studies provide material which suggests that climate change,
particularly in the eastern regions of the
steppes, was among the causes of these population movements (Van Geel et al. 2004).
If we consider that there is a correspondence between the
overall distribution of haplotypes and haplogroups and past
human movements, it seems that the European or Caucasoid component
observed in the ancient Siberian sample
may originate from East European populations. Moreover,
it is likely that some mtDNA lineages were carried to
southern Siberia from the Volga–Ural region. Incidentally,
in the Wfth century BC, Herodotus mentioned transit trade
occurring in Central Asia along a route that stretched from
the Urals in the west to the Altai and the Minusinsk Basin
in the east (Hemphill and Mallory 2004). In Altai, the presence
of the R1a1 haplogroup in the middle of the fifth century BC is
confirmed by the sample SEB 96K2 of Ricaut
et al. (2004) which was found to belong to this Y-haplogroup.
The boundary of the eastern European influence
seems to be fixed at the peri-Baikal area since no R1a1
haplogroup was found in the Xiongnu specimens of the
Northern border of Mongolia.
According to the “Kurgan hypothesis” of Marija Gimbutas,
nomadic peoples of the Volga steppe region, assumed
to speak a Proto-Indo-European language, infiltrated
Europe in three waves between 4400 and 2800 BC. Around
4400 BC, Kurgan people from the lower Dnieper and lower
Volga regions began moving along the Black Sea littoral
into the Danube Basin. They migrated in the Central Balkans
and further into Central Europe. During the middle of
the fourth millennium BC, the Kurgan culture in the North
Pontic Region continued to develop. People travelled
across western Ukraine north of the Carpathian Mountains
to Poland and Central Germany. They also moved southwest into
eastern Romania. Shortly after 3000 BC, the third
Kurgan wave (Yamna people), originating once more from
the Volga steppe, spread from Central Europe to Northwest
Germany, the east Baltic area, southern Scandinavia, the
upper Dnieper basin and Central Russia. These three waves
of migrations might explain the distribution of mtDNA and
Y-chromosome lineages observed in the present work
(Figs. 2, 3)