|
|
Post by Admin on May 18, 2022 19:08:27 GMT
The next mountain
For now the Cobra Cave tooth’s strongest Denisovan ties come from its location and its resemblance to the molar of the Xiahe mandible. While the Laotian molar is somewhat similar to those of Neanderthals, that species has never been found as far east as Laos, and genetic data show that Denisovans probably lived in Southeast Asia.
“Everything fits with what we would expect for a Denisovan lower molar,” says Bence Viola, a paleoanthropologist at Canada’s University of Toronto who wasn’t involved with the study.
Piecing together the mysterious hominins’ anatomy has been a persistent challenge because, at least for now, Denisovan fossils are so scarce. The fact that the newfound tooth is a lower molar makes confirmation even more difficult, since only the Xiahe mandible contains lower molars firmly identified as Denisovan. Without support from DNA or additional proteins, “it’s very difficult to say anything conclusive,” says Aida Gomez-Robles, a paleoanthropologist at University College London who wasn’t part of the study team.
Yet many more Denisovans might be hiding under scientists’ noses—or in cave ceilings above their heads. A dazzling array of hominin fossils have been found across Asia, many of which have been assigned to a vague catch-all group known as “archaic Homo.” In recent years, studies have suggested that some of these hominins could be Denisovans, or at least close relatives.
“Most likely we’ve been looking at Denisovans in museums and … fossil institutions for a very long time, but we haven't known what to call them,” Shackelford says.
Researchers also have more studies planned. According to Zanolli, the team is analyzing the oxygen and carbon chemistry of the tooth’s enamel. Such studies could hint at the climate in which the Denisovan girl lived, as well as what she was eating as the molar formed.
For Shackelford, one of the study’s most exciting implications is the sheer number of discoveries that lie in wait among Laos’s cave-riddled peaks. “We’ve been working there for more than 10 years,” she says, “and we still haven’t made it off the first mountain.”
|
|
|
|
Post by Admin on May 19, 2022 19:06:42 GMT
A Middle Pleistocene Denisovan molar from the Annamite Chain of northern Laos
Fabrice Demeter, Clément Zanolli, …Laura Shackelford
Nature Communications volume 13, Article number: 2557 (2022)
Abstract
The Pleistocene presence of the genus Homo in continental Southeast Asia is primarily evidenced by a sparse stone tool record and rare human remains. Here we report a Middle Pleistocene hominin specimen from Laos, with the discovery of a molar from the Tam Ngu Hao 2 (Cobra Cave) limestone cave in the Annamite Mountains. The age of the fossil-bearing breccia ranges between 164–131 kyr, based on the Bayesian modelling of luminescence dating of the sedimentary matrix from which it was recovered, U-series dating of an overlying flowstone, and U-series–ESR dating of associated faunal teeth. Analyses of the internal structure of the molar in tandem with palaeoproteomic analyses of the enamel indicate that the tooth derives from a young, likely female, Homo individual. The close morphological affinities with the Xiahe specimen from China indicate that they belong to the same taxon and that Tam Ngu Hao 2 most likely represents a Denisovan.
Introduction
From the Early to Late Pleistocene, the presence of Homo erectus is well documented in Asia, notably in China and Indonesia1,2,3. However, the taxonomic attribution of most Asian late Middle Pleistocene Homo specimens remains a matter of contention4,5,6,7. The recent description and analysis of the Harbin cranium from China has reignited this debate by suggesting its attribution to a new species named Homo longi8, but this new taxonomic attribution remains highly debated. In fact, the Harbin cranium shows close morphological similarities with other late Middle to early Late Pleistocene Asian Homo specimens from Dali, Xujiayao, Xuchang and Hualongdong, whose taxonomy remains unclear4,9,10. These fossils are considered to belong to a different taxon than H. erectus and are often grouped under the generic label ‘archaic humans’9,10. Due to the combination of features they exhibit, including Neanderthal-like traits, it has been suggested that they belong to an Asian sister taxon of Neanderthals, the Denisovans, even if this attribution remains under debate5,11,12. The small number of fossils currently securely attributed to the Denisovans (Denisova 2, a lower left molar; Denisova 3, a distal manual phalanx; Denisova 4, an upper left M3; Denisova 8, an upper molar; and the Xiahe mandible)13,14,15,16 prohibits a clear morphological picture of their overall morphology. The geographic distribution of the Denisovans also remains debated. Modern Papuans, Aboriginal Australians, Oceanic/Melanesian, Philippine Ayta groups and, to a much lesser extent, mainland Southeast Asian populations, retain a Denisovan genetic legacy14,17,18,19. Combined paleoproteomic and morphometric analyses recently suggested that the Middle Pleistocene Xiahe mandible from Baishiya Karst Cave belonged to a Denisovan, extending the known range of this group onto the Tibetan Plateau15. However, there is still no fossil evidence explaining the Denisovans genetic imprint on modern southeast Asian populations and—due to the paucity of the Middle Pleistocene fossil record—it is still unknown whether one or more human lineages (co)existed in continental southern Asia. We present here to the best of our knowledge the first unambiguous Middle Pleistocene Homo specimen from mainland southeast Asia and discuss its taxonomic attribution and implications for human evolution in the region.
In December 2018, a hominin permanent lower molar was recovered from a breccia block at Tam Ngu Hao 2 (Cobra Cave), Huà Pan province, Laos (20°12′41.5′′N, 103°24′32.2′′E, altitude 1,116 m; Fig. 1, Supplementary Fig. 1). The tower karst in which the cave was formed is positioned on the south-eastern side of P’ou Loi Mountain with an entrance located 34 m above the alluvial plain (Fig. 1a, Supplementary Fig. 1). The site was discovered during a survey of the area around Tam Pà Ling, where early Homo sapiens fossils have previously been recovered20,21,22. The tooth (TNH2-1) is a mandibular left permanent molar crown germ (Fig. 2a–f; Supplementary Fig. 2), and the absence of occlusal and interproximal wear combined with the incipient root formation suggests that the tooth was unerupted at the time of the individual’s death. The morphology of the tooth is compatible with an attribution to either a first or a second lower molar (Methods-Detailed morphological analysis of the tooth). In either case, considering the early maturational stage of the root, this tooth belonged to a juvenile individual corresponding to an age ranging from 3.5 to 8.5 years following modern developmental standards23.
|
|
|
|
Post by Admin on May 19, 2022 19:50:20 GMT
Fig. 1: Geomorphological context and stratigraphy of TNH2. 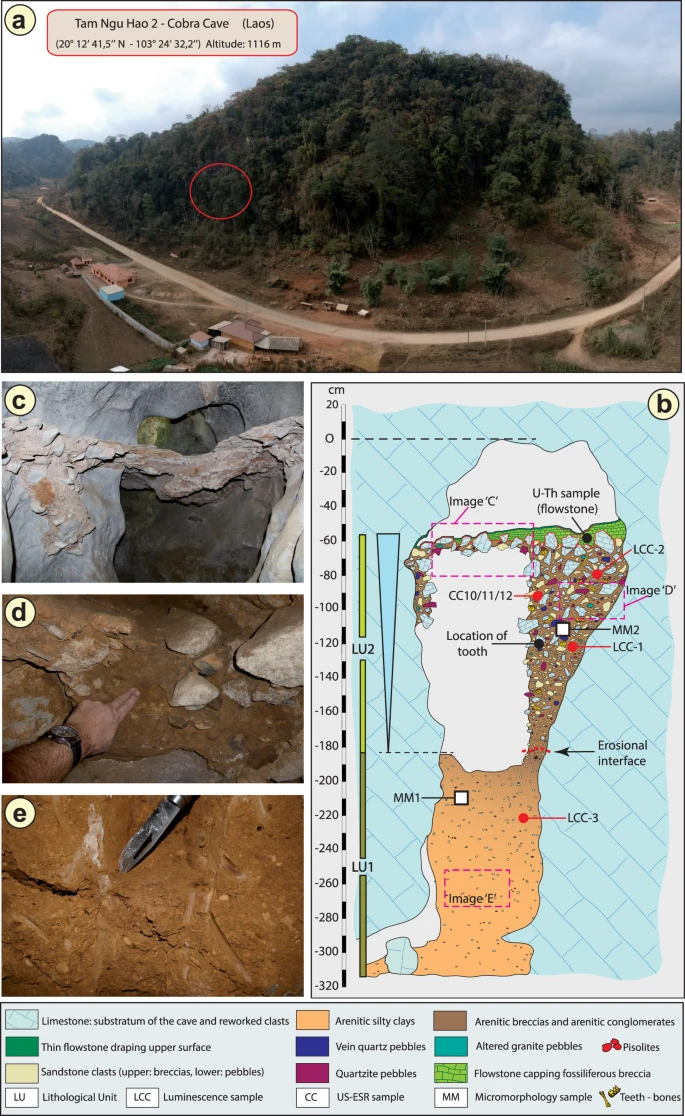 a Aerial view of the site. The red circle indicates the entrance of Tam Ngu Hao 2 cave. b Stratigraphy and sampling locations of the infilling of the cave, showing Lithological Unit 1 and 2 (LU1 and LU2) with the erosional interface between these layers indicated by a dashed red line; Micromorphological (microstratigraphic) samples (MM1 and MM2) are also shown. Encircled numbers denote approximate positions of photographs in c, d and e. c View of the flowstone capping the upper remaining part of LU2. d Detail of the arenitic breccia/conglomerate of LU2. e Detail of the arenitic silty clay of LU1. Fig. 2: Views of the TNH2-1 specimen. 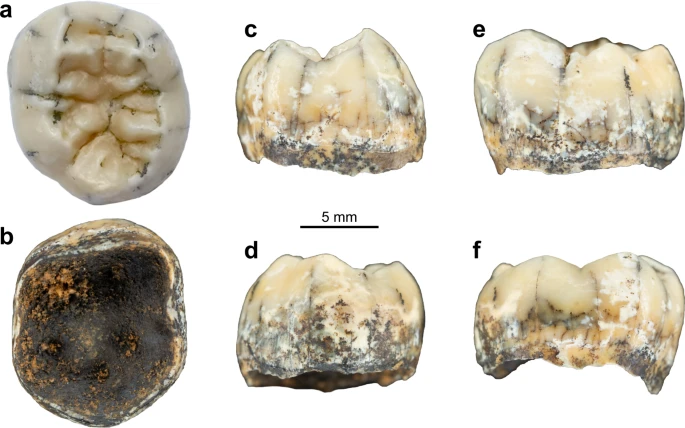 TNH2-1 in occlusal (a), inferior (b), mesial (c), distal (d), buccal (e) and lingual (f) views. In this work to best document THN2-1, morphological description and comparative analyses are performed. We also develop a specific sampling protocol that allows us to sample for palaeoproteomic and future isotopic analyses while preserving the whole occlusal surface morphology of the crown. Sampling for these destructive analyses takes place after microCT analysis of the entire tooth, ensuring full morphological data were saved. No additional sampling for ancient DNA analyses is performed at this stage given the old age of the specimen and the tropical conditions under which the sediment and fossils were deposited. The invasive sampling strategy to collect dental tissues for molecular analyses only focuses on the distal part of the inferior aspect of the crown, keeping the mesial portion of the crown intact. |
|
|
|
Post by Admin on May 20, 2022 19:29:07 GMT
Results
Context and dating
The geological setting, stratigraphy and micromorphology of the sediment sequence were analysed to obtain a comprehensive, multi-scalar assessment of the depositional context and taphonomic history of the fossils recovered from the cave (Supplementary Information, Geology). The partially eroded sediments that infill the studied entrance passage comprise a lower and an upper facies representing two phases of sediment accumulation separated by an erosional surface and an unknown period of time (Fig. 1b). The lower facies (Lithological Unit 1, LU1) is weakly cemented and forms an arenitic silty clay deposit that is devoid of fossils (Fig. 1e). The upper, fossiliferous facies (Lithological Unit 2, LU2) is well cemented and coarse grained, containing intrakarstic angular limestone clasts and extrakarstic rounded pebbles, forming a very hard breccia/conglomerate layer from which skeletal elements—and in particular, teeth—were recovered in high frequencies (Fig. 1d). The change in lithology between the two facies most likely reflects a reconfiguration of the karstic hydrological system as would be associated with a major flood, eroding space in LU1 onto which the sediments of LU2 were unconformably overlain. The sediments of LU2 are laterally contiguous and densely packed throughout the exposure excavated for this study, precluding major reworking of material and confirming the stratigraphic context of the fossils contained within, including the hominin tooth (see detailed observations described in Methods). The upper facies (LU2) is draped with two carbonate flowstones, indicating a final change in hydrology and the passage of surface water out of the cave and the precipitation of laminar speleothem (Fig. 1c).
Three bovid teeth (TNH2-10/CC10, TNH2-11/CC11, TNH2-12/CC12) recovered from the upper fossil-bearing breccia (LU2) were directly dated using coupled uranium series and electron spin resonance (US-ESR), providing a weighted mean age estimate of 151 ± 37 thousand years ago (kyr) (2-sigma) (Fig. 1b; Supplementary Tables 1, 2) and an age range of 188–117 kyr. Two large blocks of breccia (LCC1 and LCC2) from LU2 (upper) and one block of the silty clay unit (LCC3) from LU1 (lower) were removed for luminescence dating (Fig. 1b). These samples produced coeval age estimates of 143 ± 24 kyr (LCC1) and 133 ± 19 kyr (LCC2) for the deposition of the LU2 breccia and 248 ± 31 kyr (LCC3) for the underlying LU1 silty clay deposit (Supplementary Table 3). These ages are in stratigraphic agreement with the age of the overlying flowstone (CCF1), which was precipitated earlier than 104 ± 27 kyr based on the weighted mean of U-series age estimates on four separate sub-samples of flowstone carbonate (Supplementary Table 4). Bayesian modelling was performed on all independent age estimates to determine an overall geochronological framework for the site and tooth (Methods and Supplementary Fig. 3). The fossiliferous breccia including the tooth was deposited between 164 and 131 kyr (at 68% confidence limit).
Fauna
The Tam Ngu Hao 2 faunal assemblage comprises 186 identified dentognathic specimens (NISP) dominated by isolated teeth of large mammals, including several megaherbivores (Supplementary Table 5, Supplementary Data 2). Their analyses reveal typical taphonomic pathways of assemblages from karstic systems in terms of representation of specimens and types of damage. Due to the energy associated with the deposition of LU2, only teeth of large mammals are present in the assemblage, and we note the absence of small and light teeth of any microvertebrates. Moreover, most teeth are gnawed by porcupines, known to be a major accumulator agent in the region24. Therefore, the poor preservation of specimens precludes identification to the species level for most of the recorded taxa. The fauna bears close affinities to those known from the late Middle Pleistocene of southern China and northern Indochina and, to a lesser extent, Java, which is consistent with the sedimentary chronology of the site. It can be assigned to the “Stegodon-Ailuropoda faunal complex”25,26,27,28. We note the absence of Neogene taxa that persist in the Early Pleistocene and that of two key-species, Pachycrocuta brevirostris and Gigantopithecus blacki, which are good indicators of pre-300 kyr faunas in the region26,27,29,30,31. The archaic Stegodon persisted in Asia most likely until the end of the Late Pleistocene32. We recovered herbivores including Tapirus, Stegodon, and Rhinocerotidae, that were adapted to canopied woodlands in the area. We also found animals such as the Bos species, small-sized Caprinae and large-sized Cervidae (possibly Rusa unicolor), which are all known to exhibit a great variability in their feeding behaviour and preferred habitats, from closed and intermediate forests to open grassland33.
|
|
|
|
Post by Admin on May 21, 2022 18:50:00 GMT
Ancient protein analyses
The enamel from the TNH2-1 tooth specimen was analysed using nanoLC-MS/MS and the recently developed approach for ancient enamel proteomes34. The TNH2-1 proteome is composed of a common set of enamel-specific proteins, all of which have previously been observed in Pleistocene enamel proteomes34,35,36 (Supplementary Table 6). The enamel proteome has elevated levels of diagenetic protein modifications (Supplementary Fig. 4a–d, Supplementary Table 7) and preserves serine (S) phosphorylation within the S-x-E motif previously observed in ancient dental enamel34,35 (Supplementary Fig. 4e). Based on proteome composition and modification, as well as the absence of peptides matching any of these proteins in our extraction and mass spectrometry blanks, we consider our proteomic data as indicative of endogenous proteins deriving from the sampled enamel.
Unfortunately, no high-confidence peptides overlapped diagnostic amino acid positions with sequence differences between H. sapiens, Denisovans, or Neanderthals, making further taxonomic assignment based on palaeoproteomics impossible. This is in line with previous research, which indicated that closely related hominin populations can be distinguished based on dentine and bone proteomes, while enamel proteomes are less informative in the context of close phylogenetic proximity35. Nevertheless, by comparing the sequences recovered from the TNH2-1 enamel proteome with that of extant hominids for which protein sequences are available, we find that the specimen belongs to a member of the genus Homo (Supplementary Table 8).
Supplementary Table 8. Informative amino acid positions covered in the TNH2-1 proteome.
For each hominid genus, the amino acid observed in the reference sequences is indicated, as is the
amino acid observed in the TNH2-1 enamel proteome.
Protein Human ref. accession Amino acid position* Pongo sp. Gorilla sp. Pan sp Homo sp. Amino
acid in TNH2 Consequence
ALB P02768 49 I V V I I Not Gorilla,
not Pan
AHSG P02765 21 H H R H H Not Pan
AHSG P02765 23 P P L P P Not Pan
AMBN Q9NP70 270 G E E E E Not Pongo
ENAM Q9NRM1 64 N N T N N Not Pan
ENAM Q9NRM1 137 Q R R R R Not Pongo
ENAM Q9NRM1 147 A P P P P Not Pongo
ENAM Q9NRM1 190 I I M I I Not Pan
AMELX Q99217-3 126 T I I I I Not Pongo
AMELX Q99217-3 134 L P P P P Not Pongo
The absence of peptides specific to male-diagnostic amelogenin Y (AMELY) suggests that either the sampled molar was from a female individual or that AMELY-specific peptides were not observed due to degradation beyond the limit of detection of the instrument.
|
|

