|
|
Post by Admin on Oct 5, 2022 7:39:31 GMT
Fig. 5: Population structure of present-day Britain and Ireland. 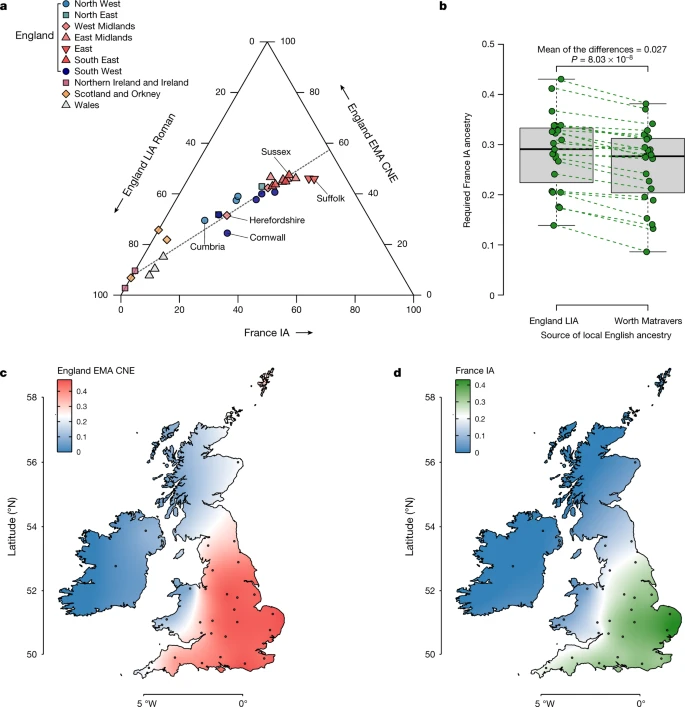 a, Ternary plot of present-day British–Irish populations as a three-way admixture between late Iron Age and Roman England (England LIA Roman) (n = 32), France IA (n = 26) and England EMA CNE (n = 109). b, Boxplot comparison of France IA ancestry proportions in 23 English PoBI sampling regions using either England LIA Roman (n = 32) or Worth Matravers (n = 16) as source for local British ancestry in qpAdm. The P value obtained from a two-sided paired Student’s t-test is shown. The bounds of the box represent the 25th and 75th percentile, the centre represents the median, and the whiskers represent the minimum and maximum values in the data. Dashed lines connect points from the same region. c, Geographical distribution of the England EMA CNE, ancestries based on the interpolation of 31 present-day population estimates. The coordinates of the sample collection districts approximate the centroids of the averaged birthplaces of the grandparents. d, Same as c, but for France IA. Our three-way population model for present-day England supports a view of post-Roman English genetic history as punctuated by gene flow processes from at least two major sources: first, the attested arrival of CNE ancestry during the Early Middle Ages from northern Germany, the Netherlands and Denmark, and second, the arrival of ancestry related to France IA. Although we cannot precisely date the order of those arrivals, at least substantial amounts of France IA-related ancestry seem to be absent in northern and eastern England during the Early Middle Ages and therefore must have arrived there subsequently. In other parts of England, however, it may have entered together with CNE ancestry or even earlier. Notably in southern England, namely, Eastry, Apple Down and Rookery Hill, several early medieval individuals already exhibit France IA-related ancestry, which probably results, at least in part, from localized mobility between the south of England and the Frankish areas of Europe during the Early Middle Ages (Extended Data Fig. 8a). Indeed, Frankish material culture is evident in these regions, particularly in Kent and Sussex58,59,60. Admixture from this second source is, therefore, unlikely to have resulted from a single discrete wave. More plausibly, it resulted from pulses of immigration or continuous gene flow between eastern England and its neighbouring regions. www.nature.com/articles/s41586-022-05247-2 |
|
|
|
Post by Admin on Oct 7, 2022 4:35:04 GMT
Discussion
The ‘Anglo-Saxon settlement’ is among the most intensely debated topics in British history, but much of the discussion remains anchored to the contents of Bede’s Ecclesiastical History and the Anglo-Saxon Chronicle18. These early writings defined the settlement as a single event, or a series of events, tied to the immediate aftermath of the Roman administration in the fifth to sixth century. In the archaeological and historical debate, this has been described as happening to varying degrees; as the Adventus Saxonum (a folk migration of named Germanic tribes), an invasion or the movement of a limited number of elite male migrants18,61. To this day, little agreement has been reached over the scale of migration, the mode of interaction between locals and newcomers, or how the transformation of the social, material, and linguistic or religious spheres was achieved. Here we provide strong evidence of large-scale early medieval migration across the North Sea zone and extend its temporal scope. In particular, we show that these migrations started earlier than previously assumed, as evidenced by individuals with CNE ancestry from later Roman contexts, and continued throughout the middle Anglo-Saxon period. Our results from middle Saxon sites such as Sedgeford push the estimated dates of arrival of CNE ancestry to as late as the eight century and merge these events with interpersonal mobility from Sweden and other Scandinavian regions during the later Viking invasion and settlement. Together, these migrations appear to be part of a continuous movement of people from across the North Sea to Britain from the later Roman period into the eleventh century CE.
Our results overwhelmingly support the view that the formation of early medieval society in England was not simply the result of a small elite migration18,61, but that mass migration from afar must also have had a substantial role. We identified numerous individuals with only continental ancestry, suggesting that many of them were migrants themselves or were their unadmixed descendants. Both the lack of genetic evidence for male sex bias, and the correlation between ancestry and archaeological features, point to women being an important factor in this migration. Although men with migrant and local ancestry were buried in similar ways, women with migrant ancestries were more often found with grave goods than women with local ancestry. This could point to social stratification, or plausibly might simply reflect the degree to which women of local ancestry were integrated into the emerging CNE families. It is clear, however, that these social differences are subtle, given that we did not find evidence for this pattern in male burials, and that we found significant regional and site-level differences. Previous hypotheses about the social mechanisms in this migration have included partial social segregation62, elite migration18,61, substantial population replacement34 or no migration at all1,22. Our combined genetic and archaeological analysis point to a complex, regionally contingent migration with partial integration that was probably dependent on the fortunes of specific families and their individual members.
In present-day Britain, we saw substantial northern continental ancestry, albeit at a lower level than during the early medieval period, pointing to a lasting demographic impact of the ‘Anglo-Saxon’ migrations. Specifically, in early medieval western England, Wales and Scotland, and more generally in England during the Norman period, further aDNA sampling may clarify how CNE ancestry spread and was subsequently diluted. Beyond the substantial early medieval immigration of northwestern continental European people found here, we have also identified a second major source of continental ancestry in modern Britain from sources more to the European south and west. This second ancestry component is already evident in our early medieval samples. In Southeast England specifically, individuals at several sites show ancestry whose closest match is in modern-day western Germany, Belgium and/or France, which matches the Frankish connections seen in the archaeological record for these regions. Our data and analyses indicate that this second genetic introgression continued further into the Middle Ages and potentially beyond.
|
|
|
|
Post by Admin on Mar 7, 2023 18:42:35 GMT
Dual ancestries and ecologies of the Late Glacial Palaeolithic in Britain Abstract Genetic investigations of Upper Palaeolithic Europe have revealed a complex and transformative history of human population movements and ancestries, with evidence of several instances of genetic change across the European continent in the period following the Last Glacial Maximum (LGM). Concurrent with these genetic shifts, the post-LGM period is characterized by a series of significant climatic changes, population expansions and cultural diversification. Britain lies at the extreme northwest corner of post-LGM expansion and its earliest Late Glacial human occupation remains unclear. Here we present genetic data from Palaeolithic human individuals in the United Kingdom and the oldest human DNA thus far obtained from Britain or Ireland. We determine that a Late Upper Palaeolithic individual from Gough's Cave probably traced all its ancestry to Magdalenian-associated individuals closely related to those from sites such as El Mirón Cave, Spain, and Troisième Caverne in Goyet, Belgium. However, an individual from Kendrick's Cave shows no evidence of having ancestry related to the Gough’s Cave individual. Instead, the Kendrick’s Cave individual traces its ancestry to groups who expanded across Europe during the Late Glacial and are represented at sites such as Villabruna, Italy. Furthermore, the individuals differ not only in their genetic ancestry profiles but also in their mortuary practices and their diets and ecologies, as evidenced through stable isotope analyses. This finding mirrors patterns of dual genetic ancestry and admixture previously detected in Iberia but may suggest a more drastic genetic turnover in northwestern Europe than in the southwest. Main The climatic warming that occurred after the Last Glacial Maximum (LGM) was critical in the development of human societies and dramatically altered the distribution of faunal and floral communities in Europe. Landscapes that were uninhabited at the LGM were recolonized during the Late Glacial and the distribution and density of human populations changed markedly, alongside the emergence of substantial cultural diversification. As such, many studies have focused on relationships between population expansion, environmental change and cultural diversity in Europe in the post-LGM period (for example, refs. 1,2,3,4,5). However, details of human postglacial recolonization of Europe remain unclear due to the complex history of prehistoric migrations across the continent and the relative paucity of human remains dating to this period. In recent years, advances in sequencing technologies, combined with improved laboratory methods and bioinformatic workflows, have opened up the possibility to generate and analyse the genetic signatures of Late Pleistocene European populations. To date, a number of studies have explored the genetic makeup of the earliest modern humans in Europe, before the emergence of agriculture. These studies have revealed numerous instances of genetic shifts indicative of population expansions6,7,8,9. One of the most notable examples occurred during the Late Glacial, between the end of the LGM (~23,400 calibrated years before present (cal. BP)) and the start of the Holocene epoch (~11,700 cal. BP). This shift is reflected in the ancestries associated with the ~15,090-year-old (IntCal20) Goyet Q2 individual, Belgium, and the ~14,010-year-old (IntCal20) Villabruna individual, Italy, in post-LGM Europe. We use these individuals as shorthand for the ancestries associated with them throughout the text. ‘Goyet Q2’ ancestry9, which has previously been defined by the ~18,770-year-old (IntCal20) ‘El Mirón’ individual8 from Spain, has been identified in individuals associated with the Magdalenian culture, dating from ~20,500 to 14,000 cal. BP. This Goyet Q2/El Mirón ancestry has been suggested to represent a post-LGM expansion from southwestern European glacial refugia8. The ‘Villabruna’ ancestry, also broadly known as Western hunter gatherers or WHG, consists of individuals dated from ~14,000 to 7,000 cal. BP associated with Epigravettian, Azilian/Federmesser, Epipalaeolithic and Mesolithic cultures8. The Villabruna ancestry is also associated with the observation that from ~14,000 cal. BP, all European individuals show some level of genetic affinity to present-day Near Eastern populations8. The expansion in the geographic distribution of this ancestry also correlates with a period of rapid climate warming of the Late Glacial Interstadial (considered broadly equivalent to the onset of Greenland Interstadial 1 (GI-1), ~14,650 cal. BP) as well as cultural transitions from the Magdalenian/Late Upper Palaeolithic to the Azilian/Federmesser-Gruppen/Final Palaeolithic and has therefore been suggested to represent the movement of people into northwestern Europe after the LGM8. Interestingly, however, individuals with a mixture of Goyet Q2 and Villabruna ancestry appear in southern Europe from at least ~18,700 cal. BP—with the individual from El Mirón8 being the earliest identified thus far. The presence of individuals with admixed Goyet Q2 and Villabruna ancestry in southern Europe from the LGM onwards raises questions related to the fragmentation of populations into isolated refugia during the last Ice Age1,10,11,12. It appears that both cultural and gene flow continued across the continent—although the nature of these processes and the mechanisms involved remain unclear. However, the presence of individuals with un-admixed Goyet Q2 ancestry in northern Europe until ~14,000 cal. BP (ref. 9) also suggests some degree of sustained isolation throughout the LGM and into the Late Glacial. There is evidence of populations living in ice-marginal environments within northern Europe at the LGM and of long-distance movement of people from east to west north of the Alps, which has also been linked to the expansion of Magdalenian cultural groups13,14,15. This evidence raises suggestions of Magdalenian populations with Goyet Q2 ancestry—who appear to have been cold-adapted hunter gatherers—retreating to northern Europe, perhaps due to climatic warming and the movement of prey species such as reindeer and horse. Conversely, more southerly regions such as northern Spain and Italy, where temperate prey species such as red deer persisted throughout the LGM and Late Glacial, may have provided greater ecological opportunities for population admixture16. Britain lies at the extreme northwest corner of the post-LGM expansion. With approximately two-thirds of the landmass covered by ice at the LGM and rapid deglaciation thereafter17, substantial ecological and environmental change took place in the post-LGM landscape. As such, Britain offers a unique environmental context through which Late Upper Palaeolithic populations can be considered. By ~19,000 cal. BP the British–Irish Ice Sheet was undergoing widespread melt and by ~16,000 cal. BP ice was absent from virtually all of England and Wales17. Reindeer were present in southwest England by ~17,000 cal. BP (ref. 18) and habitats were dominated by open steppe–tundra vegetation19,20. However, detailed consideration of Late Upper Palaeolithic sites in the United Kingdom and a series of radiocarbon dating programmes suggest that there is no evidence for post-LGM human recolonization of southwestern Britain before ~15,500 cal. BP (ref. 21). As such, some regions of Britain were colonized before the rapid climate warming at the start of the Late Glacial Interstadial (~14,650 cal. BP). Accelerator mass spectrometry (AMS) dating indicates that Britain was probably recolonized at a slightly later date than adjacent regions such as the Paris Basin and the Belgian Ardennes21,22—thereby suggesting an expansion of people across the European continent1. Interestingly, the British Magdalenian (known locally as the Creswellian) appears to be very similar (both in terms of chronology and cultural expression/typology) to the Classic Hamburgian, found in the northern Netherlands and the lowlands of northern Germany and Poland22,23. However, understanding the expansion of post-LGM populations into and within the British Isles is hindered by a relative paucity of preserved archaeological remains suitable for dating22. As such, the exact nature of human occupation of Late Upper Palaeolithic Britain remains unclear and we have relatively little knowledge of the earliest postglacial populations in Britain. Whilst the genetics of Mesolithic, Neolithic and Bronze Age individuals from Britain have recently been explored24,25, no genetic data have yet been generated for British Palaeolithic individuals, due in part to the scarcity of human skeletal material available from Late Pleistocene Britain. To date, modern human skeletal remains have been recovered from only six Upper Palaeolithic sites26,27,28,29,30. Nonetheless, these rare samples are crucial for our understanding of human populations across post-LGM Europe due to Britain’s location on the most northwesterly fringe of the European continent. Mesolithic British populations have been identified genetically as WHGs (Villabruna ancestry), indicating that this genetic ancestry spread to the most northwesterly area of early Holocene Europe by at least ~10,500 cal. BP (ref. 25). What remains unclear, however, is when this ancestry first arrived in Britain and, additionally, what the genetic ancestry of Palaeolithic populations in Britain may have been. Given the previous association of Goyet Q2 ancestry with Magdalenian cultures across Europe and the similarities between the Creswellian and the Classic Hamburgian cultures, it could be hypothesized that British Late Upper Palaeolithic populations would also fall within the Goyet Q2 genetic cluster. To address these questions and expand our knowledge of the genetic makeup of Europe after the LGM, we investigate here the genetic characteristics of Late Upper Palaeolithic Britain through ancient DNA analyses of human remains from two archaeological sites in England and Wales. www.nature.com/articles/s41559-022-01883-z |
|
|
|
Post by Admin on Mar 8, 2023 19:17:27 GMT
Materials Among two of the most well-known sites from the British Late Upper Palaeolithic are Gough’s Cave and Kendrick’s Cave. The site of Gough’s Cave is part of a large cave system situated in Cheddar Gorge in Somerset, southwest England (Fig. 1). It is particularly well-known due to its lithic and faunal assemblages being amongst the largest of any British Palaeolithic cave site investigated thus far21,22. The Gough’s Cave lithic assemblage is of mixed origin, containing both late Magdalenian and early Federmesser-Gruppen technologies and the recorded start of occupation at the site coincides with the beginning of the Interstadial period (GI-1e), when temperatures rapidly increased21,22. As well as the Mesolithic-dated ‘Cheddar Man’ skeleton, the remains of at least six Late Palaeolithic human individuals (a child, two adolescents and three adults)31 have been recovered from the site, two of which have previously been directly radiocarbon dated. The skeletal remains have been shown to exhibit considerable humanly induced modification that can be attributed to cannibalistic practices and the production of ‘skull-cups’31,32,33. Previous analysis of radiocarbon dates from the human remains, as well as humanly modified faunal material, constrain the start of the Late Upper Palaeolithic occupation of the cave to 14,840–14,680 cal. BP (68.2% probability, IntCal09) and occupation lasted little more than 200 years21,34. We targeted one human temporal bone from Gough’s Cave (PV M 96544 (excavation numbers GC 86 (55) and GC 87 (60)) for ancient DNA (aDNA) analysis. The sample derives from the petrous part of the temporal bone (GC 87 (60)) and was recovered from a directly dated context along with other AMS dated human remains (Supplementary Information). Fig. 1: Location, genetic ancestry and AMS date of individuals discussed within the text. 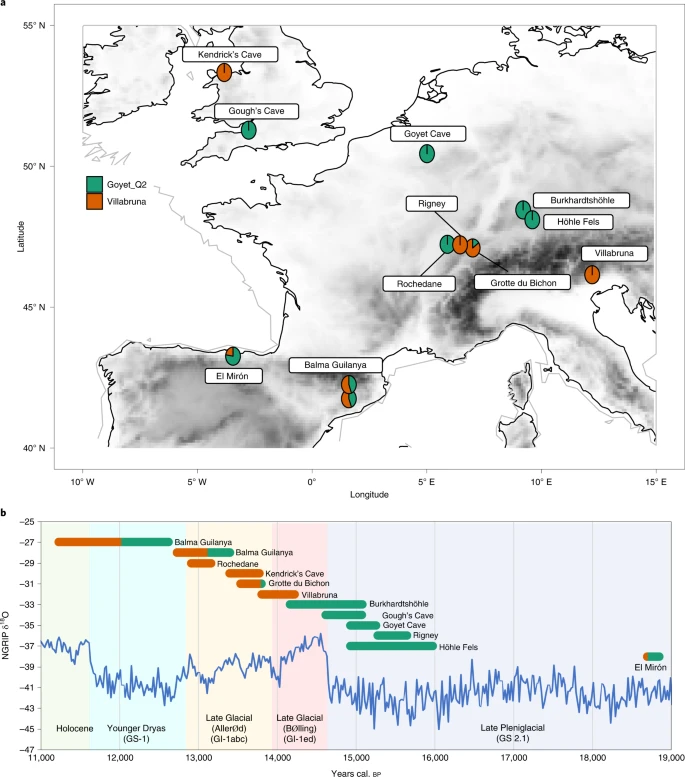 a, Map indicating the location of Pleistocene sites discussed within the text and the genetic ancestry of individuals analysed from them. b, NGRIP ice core δ18O values and INTIMATE event stratigraphy (North Greenland Ice Core Project members 2004; ref. 42), genetic ancestry and date of individuals (IntCal20, 95% confidence interval of calibrated radiocarbon dates for directly dated humans). The Gough’s Cave and Höhle Fels specimens are not directly dated and therefore for Höhle Fels specimen the age range shown is the 95% confidence interval of calibrated radiocarbon dates from bones recovered from the same area/context. For the Gough’s Cave specimen, the age range shown is the Bayesian modelled site occupation start and end dates based on AMS dating of the human remains and humanly modified faunal remains (Supplementary Table 7). Kendrick’s Cave is located on Great Orme’s Head, a limestone massif in Llandudno, North Wales (Fig. 1). The site is known to have been used by people during the early part of the Late Glacial Interstadial (GI-1e) and has been associated with Magdalenian technologies due to the recovery of a proximal portion of a broken blade with en éperon butt preparation within the cave and a cut-marked bovine bone dated to ~14,500 cal. BP (OxA-17726, 12,310 ± 50 14C BP)22,35. A decorated horse mandible dated to ~12,900 cal. BP (OxA-X-2185-26, 11,050 ± 90 14C BP) shows that the site continued to be used later into the Late Glacial Interstadial. Beads made from the teeth of brown bear, aurochs and red deer, along with the remains of a minimum of four human individuals (three adults and one child) were also recovered from the site36,37. It has been argued that these incised and perforated artefacts share stylistic similarities with continental Late Palaeolithic art (Federmesser-Gruppen: the ‘penknife point’ culture)38. Five direct radiocarbon dates have been obtained from the human remains, providing dates from 11,990 to 11,905 14C BP. However, only one of these dates included ultrafiltration in the pretreatment procedure21. Furthermore, the diet of these individuals has not previously been considered when calibrating these dates, which is necessary due to a marine and/or freshwater component in their diet30. Here, we redated four of the human bones and incorporated dietary information into the radiocarbon calibration (Results, Discussion and Supplementary Information). One human from Kendrick’s Cave (Kendricks_074) was also targeted for aDNA analysis. This sample derives from a mandibular first molar (M1) and the mandible from which this tooth derives has previously been AMS dated (with ultrafiltration) to 11,905 ± 50 14C BP (OxA-17089). The two sites, although chronologically close, show differences in funerary behaviour. Kendrick’s Cave has typically been interpreted as a burial site, in part due to the lack of faunal remains indicating food-processing activities or refuse at the site30,36. There is also no evidence of human modification on the Kendrick’s Cave human remains. In contrast, the human remains from Gough’s Cave were recovered from the same archaeological context as the faunal remains and both show significant human modification32. The human skeletal assemblage consists of scattered, highly fragmentary postcranial bones and relatively complete cranial vaults—some of which have, as already mentioned, been modified into skull-cups which has been interpreted as evidence for ritualistic cannibalism31,33,39. |
|
|
|
Post by Admin on Mar 9, 2023 18:35:29 GMT
Results AMS dating and stable isotopic analysis Four new ultrafiltered radiocarbon determinations were obtained from the Kendrick’s Cave human skeletal remains, which range from 11,990 ± 50 14C BP (OxA-V-2794-27C) to 11,830 ± 50 14C BP (OxA-V-2794-34C). These dates are all statistically indistinguishable from one another and from the previously published ultrafiltered date from the human mandible (OxA-17089, 11,905 ± 50 14C BP) (error weighted mean = 11,932 ± 23 14C BP, T = 6.8, d.f. = 4, P < 0.005). The dates fall within the range of the non-ultrafiltered previously published from the site (12,090 ± 90 14C BP (OxA-6144) to 11,760 ± 90 14C BP (OxA-7002)) but provide a more constrained date range, adding support to the interpretation that Kendrick’s Cave may have been used as a burial site. New δ13C and δ15N data obtained for these four samples were consistent with previous results attained. Previous isotopic analysis of the human remains suggested a diet which included marine and/or freshwater resources30,40, meaning the influence of reservoir effects on the AMS dates must be considered. A Bayesian mixing model was constructed to calculate the proportional influences of different food sources, using the programme FRUITS41, incorporating new and existing stable isotope data (Supplementary Information). A Bayesian model using OxCal (v.4.4) and the Mix_Curves function was applied to the Kendrick’s Cave humans and culturally modified faunal radiocarbon dates (Supplementary Information and Supplementary Table 7). This gives a boundary start date for human activity at the site of 16,410–14,070 cal. BP and a boundary end date of 13,730–13,140 cal. BP (95% confidence). When only the dates for the human remains are used in the model, however, the boundary start date is 14,100–13,460 cal. BP (95% confidence). The human individual used for aDNA analysis here (Kendricks_074) has a modelled date of 13,770–13,390 cal. BP (95% confidence, OxA-17089). However, as the stable isotope data from the Kendrick’s Cave humans has led to conflicting dietary interpretations regarding the proportions of marine and freshwater protein in the diet30,40, modelled dates should be treated with some caution until additional dietary information is available via, for example, compound specific isotope analysis or δ34S. Additionally, a Bayesian modelling approach using OxCal (v.4.4) and the IntCal20 calibration curve was applied to the published radiocarbon dates of the Gough’s Cave humans and humanly modified fauna (Supplementary Information and Supplementary Table 6). The results of this give a boundary start date for the site of 15,070–14,850 cal. BP and a boundary end date of 14,960–14,610 cal. BP with a 95% confidence interval. This new calibration shifts the site occupation to being primarily before the rapid climate warming at the start of the Late Glacial Interstadial (~14,700 BP) as recorded in the Greenland ice cores (GI-1e)42, although it is possible that the end of the occupation may have occurred just after the onset of the Late Glacial Interstadial. The new AMS dates and recalibration of dates for both the Gough’s Cave site and Kendrick’s Cave individual demonstrate that although the sites are close in age, there is no overlap between the dates of the Gough’s Cave occupation and the Kendrick’s Cave humans (at 95% confidence). However, there is an overlap between the Kendrick’s Cave humanly modified bovid bone and the Gough’s Cave human occupation (at 95.4% confidence), indicating that the two sites were occupied contemporaneously. When comparing the boundary end date for the Gough’s Cave occupation to the date of the Kendrick specimen sampled for aDNA, there is at least 600 years age difference when 100% terrestrial-based diet is assumed for both. However, this is probably an underestimate as incorporating the reservoir effect for marine and/or freshwater dietary components for the Kendrick’s individual would make it younger in date and increase the age gap between the two individuals to a minimum of 840 years and potentially up to ~1,200 years (Supplementary Information). aDNA analyses We recovered 15,497 and 9,702 unique mitochondrial DNA fragments, respectively, for the Gough’s Cave and Kendrick’s Cave individuals, resulting in 53.8-fold and 34.8-fold average coverage of their mtDNA genomes. We also recovered 30,587,614 and 29,326,159 nuclear DNA fragments from Gough’s Cave and Kendrick’s Cave individuals by direct shotgun sequencing, amounting to an average of 0.53-fold and 0.48-fold genomic coverage for the two individuals, respectively. The proportion of DNA fragments mapping to the human reference genome for the Gough’s Cave and Kendrick’s Cave individuals was 23% and 18%, with an average fragment length of 62 base pairs (bp) and 63 bp, respectively. Each DNA fragment from the two individuals was seen on average between 1.95 and 2.29 times in sequencing, translating to clonality of 49% and 56%. We reconstructed complete mtDNA sequences of both individuals and determined their haplogroups using HaploGrep43 and the Phylotree database (build 17). The Gough’s Cave individual carries substitutions that define the haplogroup U8a (0.97 posterior support) and the Kendrick’s Cave individual carries substitutions that define the haplogroup U5a2 (1.0 posterior support). The U8a haplogroup has not previously been detected in British early prehistoric individuals but has been identified in Magdalenian individuals elsewhere in Europe, for example at Hohle Fels and Brillenhöhle, Germany, and the Goyet Q2 individual from Belgium8. A number of British Mesolithic individuals have previously been found to carry the U5 mt haplogroup, including one from Kent’s Cavern which has also been determined as U5a2 (ref. 25). We determined the sex of the two individuals by assessing the number of DNA fragments that align to the X and Y chromosomes44. We found that the individual from Gough’s Cave is female and the individual from Kendrick’s Cave is male (Supplementary Information). The Kendrick’s Cave skeletal material was referred to as representing ‘men’36 in original descriptions of the site but no formal osteological assessment of biological sex was undertaken. The data generated for the Gough’s and Kendrick’s Cave individuals contains 506,151 and 476,347 single nucleotide polymorphisms (SNPs) respectively, overlapping the ‘1240k’ SNP panel45,46 informative of the genetic relationships amongst ancient and present-day humans. In a principal component analysis47,48 (Supplementary Information), the Gough’s Cave individual falls close to the ~15,000-year-old Goyet Q2 individual from Belgium, whereas the Kendrick’s individual clusters with individuals with predominantly WHG-like ancestry, including British Mesolithic individuals (Fig. 2). By computing f3 statistics in the form of f3(Gough’s/Kendrick’s, ancient; Mbuti), which measures the amount of shared genetic drift between pairs of ancient individuals after their separation from an outgroup (in this case Mbuti from ref. 49), we again found that the Gough’s Cave individual shares most drift with the individuals belonging to the ~19,000–14,000-year-old Goyet Q2 genetic cluster, whereas the Kendrick’s Cave individual shares most drift with the individuals belonging to the ~14,000–7,000-year-old Villabruna genetic cluster (Fig. 3). Fig. 2: Principal component analysis (PCA) of ancient individuals. 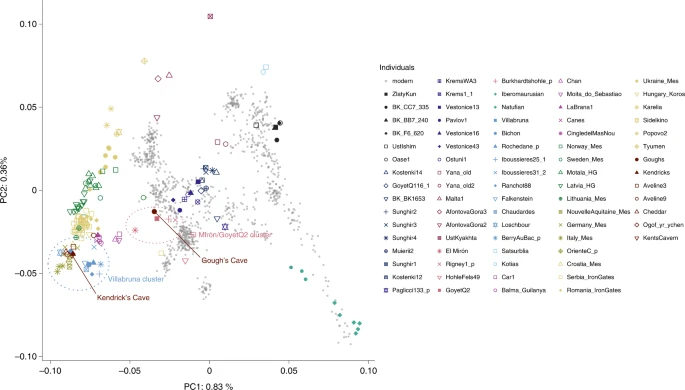 PCA of 1,087 present-day West Eurasians genotyped on 597,573 SNPs (grey points) with 168 ancient hunter gatherers (Supplementary Table 3) from West Eurasia and North Africa older than ~7,000 cal. BP and with more than 30,000 SNPs projected onto the plane. PC1, principal component one; PC2, principal component two. Fig. 3: Genetic clustering of ancient individuals, including Gough’s Cave and Kendrick’s Cave individuals, calculated as f3(ancient1, ancient2; Mbuti). 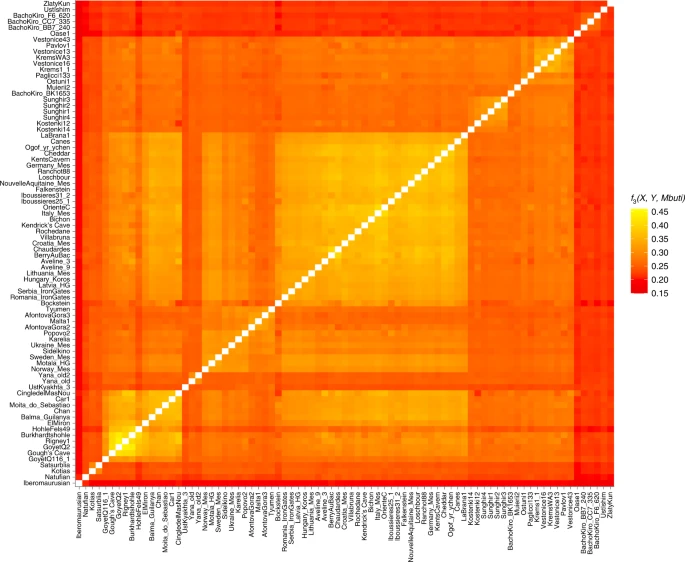 Lighter colours correspond to higher f3 values and indicate a higher shared genetic drift between pairs of ancient individuals/populations. The X-axis corresponds to ancient1 and the Y-axis to ancient2 individuals for the calculation f3(ancient 1, ancient 2; Mbuti). The order of individuals is based on the hierarchical clustering of f3 values, and ancient individuals included in this analysis are the same as those in Figure 2. Plotted f3 values were calculated using ADMIXTOOLS as implemented in admixr. We further confirmed these affinities directly with D statistics by calculating D(ancient1, ancient2; Gough’s/Kendrick’s, Mbuti). Again, we found that the Gough’s Cave individual shares significantly more alleles with the members of the Goyet Q2 genetic cluster than the Villabruna genetic cluster. In contrast, the Kendrick’s Cave individual, as well as Mesolithic individuals from Britain25, share significantly more alleles with the members of the Villabruna cluster than with the members of the Goyet Q2 cluster. We used admixture modelling with qpWave and qpAdm50,51 to explore the ancestry of the Gough’s and Kendrick’s Cave individuals in more detail. We used the Goyet Q2 and Villabruna individuals as potential source populations. We modelled the Gough’s individual as having single-source Goyet Q2 ancestry (P = 0.841) and the Kendrick’s Cave individual as having single-source Villabruna ancestry (P = 0.646). All other single-source models can be rejected (P << 0.001). Interestingly, all Mesolithic individuals from Britain, except Cheddar Man25, can also be modelled as having un-admixed Villabruna ancestry in this analysis. Cheddar Man, an individual also recovered from Gough’s Cave and dating to 10,564–9,915 cal. BP (IntCal20, 9,100 ± 100 14C BP (OxA-814)), is instead best modelled as having 84.6% (±0.5%) Villabruna-related ancestry and 15.4% (±0.5%) Goyet Q2-related ancestry (Fig. 4). Fig. 4: Modelling West Eurasian hunter-gatherer individuals (n = 26) as having a single-source or a two-source ancestry with qpAdm, using the lowest rank model. 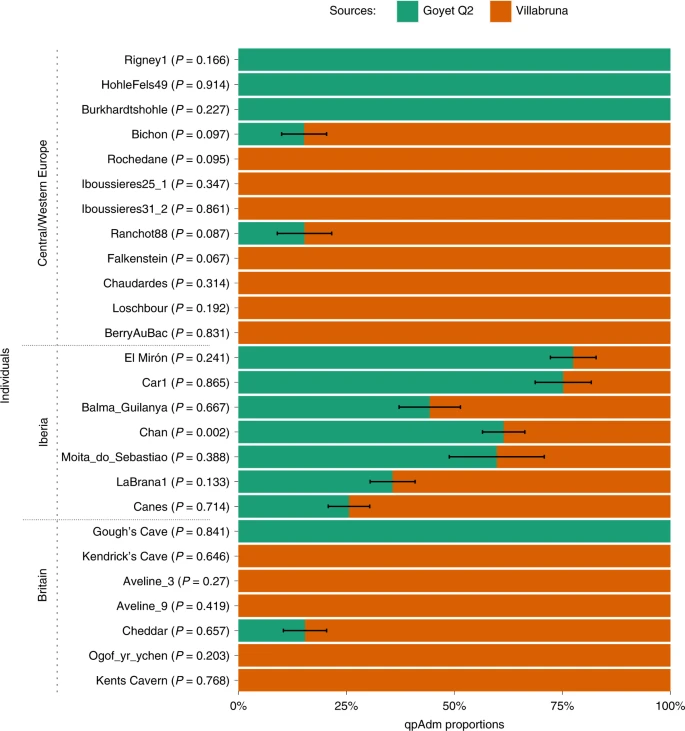 The ~15,140-year-old Goyet Q2 individual from Belgium and the ~14,060-year-old Villabruna individual from Italy were used as potential sources. Within each geographical area (Central/Western Europe, Iberia and Britain) individuals are ordered on the basis of their chronological age from oldest to youngest. Standard errors (s.e.) were calculated using weighted block jackknife and a block size of 5 Mb. Here, s.e. indicates ±1 s.e. However, it should be noted that although single-source models can explain the data from Mesolithic individuals from Britain25 and no additional model complexity is needed, two-source models of genetic ancestry of these previously published Mesolithic individuals cannot be fully rejected either. In these two-source models, Villabruna ancestry remains the predominant component for all individuals, corresponding to between 74.8% (±9.7%, Aveline9) and 93.2% (±6.7%, Ogof yr ychen) of the genetic ancestry of these individuals. In contrast, the Gough’s Cave Magdalenian individual can only be modelled as having a single-source Goyet Q2 ancestry and the Kendrick’s Cave individual can be modelled as having a single-source Villabruna ancestry, with two-source models either strongly rejected or with estimated admixture proportions outside the range 0–100%, indicative of poor fit (Supplementary Fig. 12). |
|

