

Post by Admin on Apr 5, 2019 18:40:33 GMT

Opportunities to investigate the population dynamics of early anatomically modern human (AMH) populations are rare owing to a dearth of human remains, with wide variations in ancient population size estimates from ethnographic and archaeological data (1, 2). In the absence of evidence for true contemporaneity among individuals recovered archaeologically, the population structure of foraging groups is even harder to establish. Exceptions are cases of multiple Upper Paleolithic individuals buried simultaneously or originating from sufficiently close temporal and spatial proximity that they may represent a single social group.
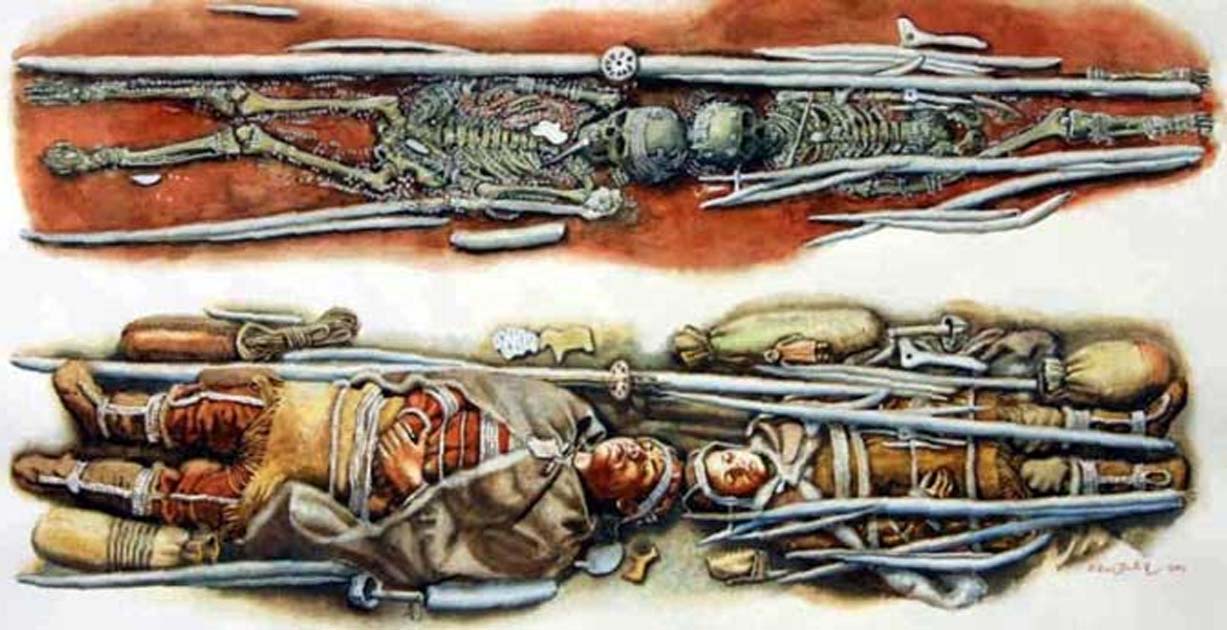
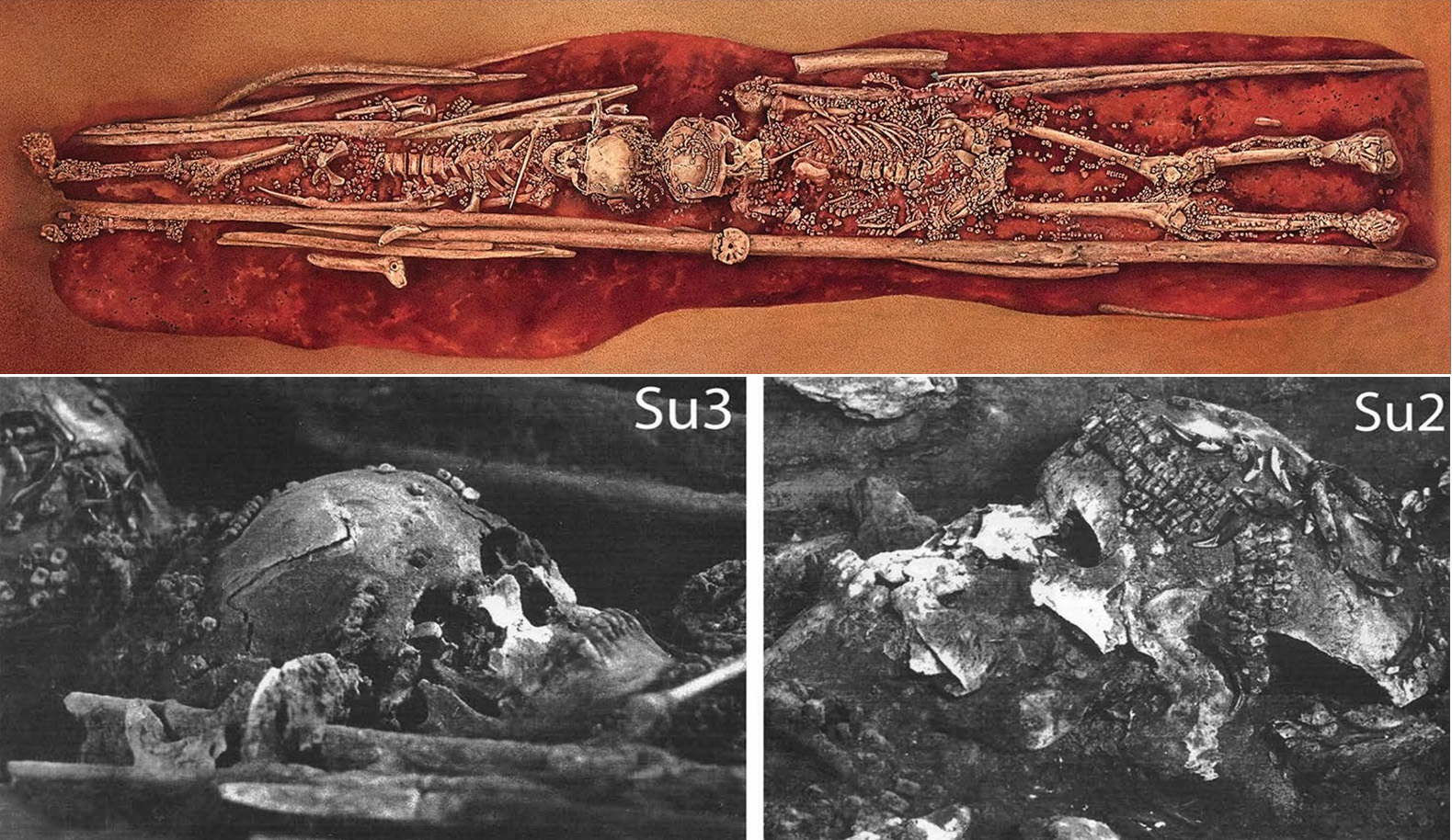
One such example of multiple burials is Sunghir, a site harboring two of the most extraordinary Upper Paleolithic burials known (3, 4) (figs. S1 and S2 and tables S1 to S4): one of an adult male [Sunghir 1 (SI)] and another of two sub-adults [Sunghir 2 and 3 (SII and SIII)], originally thought to be a boy and girl, interred head-to-head. All remains were covered in ochre and were accompanied by rich grave goods including ivory beads and spears, armbands, and carvings, as well as arctic fox canines. Adjacent to SII was the femoral diaphysis of an adult [Sunghir 4 (SIV)] that had been polished, hollowed out, and filled with red ochre. The site also yielded other less complete human remains, some of uncertain stratigraphic provenance [Sunghir 5 to 9 (SV to SIX)]. Radiocarbon analyses place the age of SI to SIV between 33,600 and 34,600 years (5, 6). The homogeneity in morphological traits (e.g., metopism) among the remains, as well as signs of possible congenital pathologies in SIII, have been interpreted as evidence of inbreeding (3). Other Upper Paleolithic individuals with reported congenital or degenerative pathologies (e.g., at Barma Grande and Dolní Věstonice) (3) reinforce the view that Upper Paleolithic groups were small and susceptible to inbreeding, possibly as extensive as what has been reported for the Altai Neandertal (7). However, genomic data available for some of those individuals (8) were of insufficient coverage to infer population sizes or inbreeding levels.

Fig. 1
Relatedness among ancient Eurasians.
Kinship coefficients and R1 ratios (4) were inferred from IBS counts. (A) Pairs of Upper Paleolithic individuals, using 1000 Genomes Phase 3 single-nucleotide polymorphism (SNP) sites. (B) Pairs of ancient Eurasians, using 1240K capture SNP sites. Within-group pairs of Sunghir are highlighted.
We screened six of the Sunghir individuals (SI to SVI) to assess DNA preservation; five of them (all but SV) yielded sufficient endogenous DNA for genome sequencing. We sequenced these genomes to an average depth of coverage ranging from 1.11× to 10.75× (figs. S3 and S4 and tables S5 and S6) and compared them to panels of modern and ancient human genomes (4). All individuals were genetically male on the basis of the fraction of Y chromosome reads (table S7), including SIII, who was previously identified as female (3). Contamination levels from X chromosome heterozygosity were low (0.33 to 0.90%; table S5), except for SVI (13.1%). Radiocarbon dates indicate that whereas SV is only slightly more recent than the other individuals, SVI is from ~900 years before the present (figs. S5 and S6 and tables S8 to S10). Together with mitochondrial DNA (mtDNA) and Y chromosome haplogroups (W3a1 and I2a1b2, respectively), these data indicate that SVI is not associated with the Upper Paleolithic burials at the site; SVI was therefore excluded from further analyses.
Analyses of mtDNA genomes place SI to SIV in haplogroup U, consistent with West Eurasian and Siberian Paleolithic and Mesolithic genomes (9) (fig. S7 and table S5). SI belongs to haplogroup U8c; the sequences for the three individuals from the double burial (SII to SIV) are identical and belong to haplogroup U2, which is closely related to the Upper Paleolithic Kostenki 12 (8) and Kostenki 14 (10) individuals. Phylogenetic analyses of the Y chromosome sequences place all Sunghir individuals in an early divergent lineage of haplogroup C1a2 (fig. S8 and tables S12 to S15). Y chromosome haplogroup C1, which is rare among contemporary Eurasians, has been found in other early European individuals, including the ~36,000-year-old Kostenki 14 (11).
We investigated the degree of relatedness among the Sunghir individuals with a method that allows relationship inferences up to the third degree but does not rely on allele frequencies (4). Surprisingly, none of them were found to be closely related (that is, third degree or closer), even though the SII to SIV individuals buried together share both mitochondrial and Y chromosome lineages (Fig. 1 and tables S16 to S23). We then inferred genomic segments that were identical by descent (IBD) and homozygous by descent (HBD) from three higher-coverage Sunghir genomes (SII to SIV) and a panel of ancient and contemporary humans (4). We compared their distributions to those inferred from whole genomes obtained using coalescent simulations (12) of randomly mating populations with varying effective population sizes (NE) (fig. S9). The distributions of HBD tracts were different between AMHs and archaic humans, indicating small effective population sizes and/or recent inbreeding in archaic individuals, particularly the Altai Neandertal (7, 13) (Fig. 2A and figs. S10 to S14).


