|
|
Post by Admin on Jun 21, 2018 18:50:15 GMT
Discussion
The advent of farming in Ireland lies at the temporal and geographical terminus of several thousand years of transition across Europe and seems to occur rapidly ∼ 3750 BC (47). The nature of this transition remains a long-standing archaeological controversy between proposals of migration by incoming farmers versus those of adoption of agriculture by indigenous Mesolithic populations (48⇓⇓–51). The Irish MN female farmer (3343–3020 cal. BC) from a Megalithic tomb in Ballynahatty near Belfast affords, to our knowledge, a first direct genetic view of the transition at Europe’s western edge. She displays predominant ancestry from early farmers that ultimately originated in migrating agriculturists from the Near East. This derivation is attested by her PCA and ADMIXTURE profile, her correlated allele frequencies with other Neolithic genomes, reflected in D statistics, and by her haplotypic affinity with modern southern Mediterranean populations such as Sardinians. Her early European farmer coefficient is estimated at ∼60%; an ancestry which is difficult to reconcile with extensive indigenous adoption of agriculture in Ireland only several hundred years earlier. She shares higher levels of genetic drift with Early and MN samples from Spain rather than those from Germany, supporting a link between the early farming cultures of Atlantic Europe and arguing for the possible passage of farming to Ireland via a southern coastal route rather than via the migrations through central Europe (2). A high affinity to Scandinavian farmer, Gok2 is more difficult to interpret as it is associated with the TRB (Funnelbeaker) culture whose origins are generally derived from Central European farming cultures but perhaps it is noteworthy that this later sample is also recovered from a Megalithic context.
From examination of the fraction of her genome, which is under ROH, she seems similar to other ancient Neolithics, suggesting that she belonged to a large outbreeding population. This analysis argues against a marked population bottleneck in her ancestry, such as might have occurred had she been descended from a small pioneering group of migrating farmers. Either a restricted colonization does not reflect the nature of the first Irish farmers or her ancestry was augmented by substantial additional Neolithic communication from elsewhere in intervening centuries.
However, like other MN genomes, Ballynahatty shows some elevation of hunter–gatherer ancestry relative to earlier Neolithic samples, suggesting admixture with surviving Mesolithic populations somewhere within the temporal and spatial trajectory between the Neolithic heartland and north east Ireland ∼3300 BC (5, 9). Further sampling will be needed to determine the local or continental origins of these hunter–gatherer ancestral components but it is interesting that she shows slightly higher affinity to a Luxembourger Mesolithic genome, than to Hungarian and Spanish equivalents.
There are several lines of evidence for discontinuity between the MN and Early Bronze Age within our genome samples from northeast Ireland. First, they occupy different regions in the PCA plot; second, there is a signal from the ADMIXTURE analysis of a Caucasus ancestral component within the Rathlin genomes, which is absent in Ballynahatty and other western MN genomes. Third, D statistics also show highly significant evidence for this eastern introgression. Finally, each Bronze Age sample (all male) exhibits the Bronze Age-associated Y chromosome lineage R1b-M269, the appearance of which has been strongly linked with Steppe incursion into Central Europe (9). Thus, it is clear that the great wave of genomic change which swept from above the Black Sea into Europe around 3000 BC washed all of the way to the northeast shore of its most westerly island. At present, the Beaker culture is the most probable archaeological vector of this Steppe ancestry into Ireland from the continent, although further sampling from Beaker burials across western Europe will be necessary to confirm this. The extent of this change, which we estimate at roughly a third of Irish Bronze Age ancestry, opens the possibility of accompanying language change, perhaps the first introduction of Indo-European language ancestral to Irish. This assertion gains some support by the relative lack of affinity of non-Indo-European speakers, Basques, to the ancient Bronze Age genomes (Fig. 3).
The high coverage of both Ballynahatty and Rathlin1 allows a sensitive test of haplotype donation to modern populations, with interesting and contrasting results. Whereas Ballynahatty shows closest affinity with the southwest Mediterranean, Rathlin1 has highest sharing with the geographically closest modern populations, a trend not seen with the other high coverage ancient genome samples. This affinity with Irish, Scottish, and Welsh (a weaker signal from modern English populations is undoubtedly due to the effects of Anglo-Saxon migrations; ref. 36) suggests a degree of continuity stretching over 4,000 y at the insular Celtic edge of Europe. Ireland shows the global maxima for frequencies of the R1b1a2a1a2c (M529) Y chromosome haplotype, lactase persistence, and the C282Y hemochromatosis allele. Our data show that these genotypes had arrived by the time of the Irish Bronze Age. Hemochromatosis, known as the “Celtic” disease (44), is a variably penetrant recessive disorder, potentially fatal, due to excessive retention of dietary iron. It has been suggested that the high frequencies of both C282Y and H63D are due to heterozygote advantage related to nutritional advantage in Neolithic iron-poor diets (52), mitigation of celiac disease (53), and increased resistance to parasitic infection (54). To our knowledge, our data provide the first evidence for known Mendelian disease alleles in ancient genomes and mark the associations of both hemochromatosis alleles with the island of Ireland as ancient.
PNAS 2016 January, 113 (2) 368-373
|
|
|
|
Post by Admin on Sept 23, 2018 18:33:49 GMT
 The Journal of Human Genetics has published an article, analyzing mitochondrial DNA from the burial mounds of the Eneolithic and Bronze Age in the Northern Black Sea Region. The first author of the article was prof. Alexey Nikitin (Grand Valley State University), then co-authored by Ukrainian archeologists and geneticists (Institute of Archeology of the National Academy of Sciences of Ukraine and Odessa National University). The region of the Northern Black Sea Region attracts close attention of both archaeologists and paleogeneists. It is associated with the migration of steppe nomads (presumably, a pit culture), which, according to some authoritative experts, brought not only radical cultural innovations, but also an important contribution to the formation of the gene pool of Europe in the Bronze Age. The authors of the article note that the west of the northern Black Sea region - namely from there they were taken samples analyzed - in the era of early metal (5-2 millennium BC) served as a connecting strip between the Balkan-Carpathian region (with the cultures of highly developed agriculture and metallurgy, as cucuten-trypillya and gumelnitsa) and tribes of nomadic cattle-breeders of the Black Sea-Caspian steppes. A distinctive feature of this region is burial in barrows. Probable founders of the tradition of building mounds in the Eneolithic were the steppe nomads of the Srednegorovsky (Stogovsky) cultural complex (4750-4200 BC); These burial mounds were built up and used by the representatives of the pit culture of the early Bronze Age (3200-2200 BC), later the barrows were used in catacomb culture (2700-2000 thousand years BC) and in the Babino culture or multi- ceramics (2250-1750 thousand years BC). That is, in the barrow tradition, archaeologists see cultural continuity. And geneticists are always interested in whether there is a genetic continuity.  The paper presents the results of studying mtDNA of 16 samples from five burial mounds, the location of which is indicated by the letters D, L, R, K, in the steppe zone, north of Odessa. The burial places are related to the Srednestogovsky cultural complex, to the pit culture, the catacomb culture and the Babino culture. The mounds were stratified, burials of different cultures were in different layers, this applied to all burial mounds. The authors describe some characteristics of burial mounds. Thus, the mound K1 was the highest (6.45 m in height). Four of the five burial mounds had at the base of the construction of the Eneolithic period. Eneolithic burials in the K1 and K2 burial mounds belonged to the Srednestogovsky cultural complex (representatives of this culture, obviously, were the first builders of mounds). In the mound R, the burial R3.19a-b is the oldest of all burials of the region (5450 ± 80 years old). The mound L is the only one in which the main burial belongs to the pit culture. Samples for genetic analysis were taken in consideration of their safety and cultural identity, so that different cultures were represented in the analysis. |
|
|
|
Post by Admin on Sept 24, 2018 18:23:13 GMT
 The authors describe some characteristics of burial mounds. Thus, the mound K1 was the highest (6.45 m in height). Four of the five burial mounds had at the base of the construction of the Eneolithic period. Eneolithic burials in the K1 and K2 burial mounds belonged to the Srednestogovsky cultural complex (representatives of this culture, obviously, were the first builders of mounds). In the mound R, the burial R3.19a-b is the oldest of all burials of the region (5450 ± 80 years old). The mound L is the only one in which the main burial belongs to the pit culture. Samples for genetic analysis were taken in consideration of their safety and cultural identity, so that different cultures were represented in the analysis. The authors of the article were extracted and analyzed in the laboratory of Grand Valley State University in compliance with all the conditions necessary for working with ancient DNA. From 16 samples for 14 samples, it was possible to sequester the polymorphic region (HVR-1), perform the SNP analysis and determine the mitochondrial haplogroup. 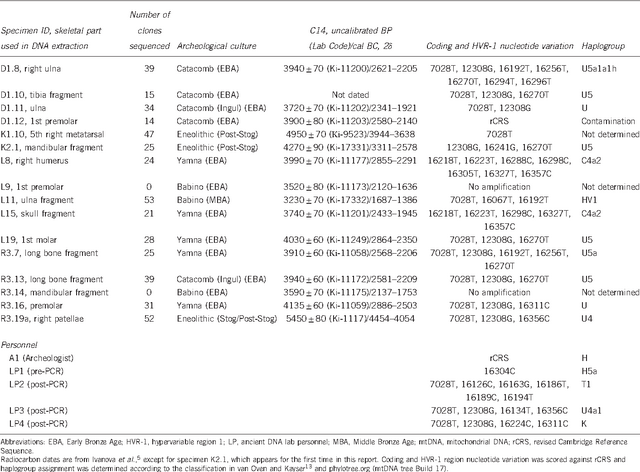 The table shows samples from the studied barrows, their dating, belonging to archeological cultures and the belonging of mtDNA to haplogroup. Of the 12 haplotypes studied, as the authors specify, 75% belonged to the West-Eurasian haplogroups of U-sub-U4 and U5. In the two samples (both from the barrow L, refer to the pit culture), an East Eurasian haplogroup C (sub-C4a2) was found. Because of the absolute predominance of mitochondrial haplogroup U in samples belonging both to the Aeneolithic (post-Stogov culture) and to the early Bronze Age (pit, babino, catacomb cultures), the authors make the main conclusion about the genetic continuity along the maternal lines of populations in the west of the North Black Sea coast during the Eneolithic - early bronze. In this case, segregation by crops was not observed in the mtDNA lines. But representatives of the Yamnoy culture along with the U line found the eastern Eurasian line S. |
|
|
|
Post by Admin on Sept 26, 2018 18:29:37 GMT
Moreover, genetic primacy can be traced to much more ancient times, since the mitochondrial haplogroup U is a haplogroup found in European hunter-gatherers of the Mesolithic, including in the north of Europe (the South Deer Island on Lake Onega). And not so long ago in the samples from the same burial ground, a mitochondrial haplogroup C was also found (this study can be read on the site ). On the graph of the main components, the authors arranged on the frequencies of haplogroups of mtDNA 39 ancient populations from different epochs (previously published data). 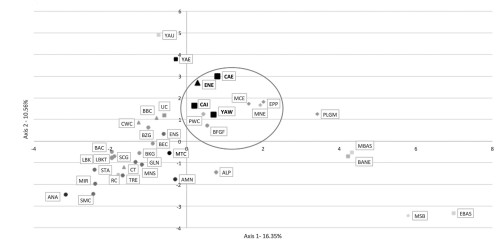 The graph of the main components is constructed from the frequencies of the mtDNA haplogroups of 39 ancient populations: Paleolith, epipaleolith, mesolite (rhombus), Neolithic (circles), Eneolith (triangles), Bronze Age (squares). The archaeological cultures of the Black Sea-Caspian region are shown in black, the cultures of Asia Minor and Anatolia are blackened by 75%, the cultures of Western and Eastern Europe are blackened by 50%, Central Europe by 35%, northern Europe by 25%, East Asia by 15% . The oval in the center indicates a cluster of the early metal era of the Northern Black Sea Coast and the European Stone Age.  It can be seen that the cultures of the steppe Eneolith (ENE), the pit culture of the west of the Northern Black Sea (YAW), the catacomb culture (CAI, CAE) and much more ancient ones, surrounded by an oval in the center, included epipaleolith of Europe (EPP), Mesolithic of Northern Europe (MNE), the Mesolithic of Central Europe (MCE). So the main conclusion of the authors of the article is about the genetic connection of the Eneolithic populations - the Bronze Age of the Northern Black Sea Coast with Mesolithic hunter-gatherers of Europe. Naturally, we are talking only about the maternal lines of inheritance - on mitochondrial DNA. Subdivisions of haplogroups U and C encompass mitochondrial DNA lineages of Eneolithic-Early Bronze Age Kurgan populations of western North Pontic steppe Alexey G Nikitin, Svetlana Ivanova, Dmytro Kiosak, Jessica Badgerow and Jeff Pashnick Abstract Prehistoric Europe experienced a marked cultural and economic shift around 4000 years ago, when the established Neolithic agriculture-based economy was replaced by herding-pastoralist industry. In recent years new data about the genetic structure of human communities living during this transition period began to emerge. At the same time, the genetic identities of the Eneolithic and Early Bronze Age (EBA) inhabitants from a prehistoric cultural crossroad in western North Pontic steppe region remain understudied. This report presents results of the investigation of maternal genetic lineages of individuals buried in kurgans constructed during the Eneolithic-EBA transition in the western part of the North Pontic Region (NPR). Mitochondrial DNA (mtDNA) lineages from the interments belonging to the Eneolithic as well as the EBA cultures such as Yamna (Pit Grave), Catacomb and Babino (Mnogovalikovaya or KMK) were examined. In the 12 successfully haplotyped specimens, 75% of mtDNA lineages consisted of west Eurasian haplogroup U and its U4 and U5 sublineages. Furthermore, we identified a subgroup of east Eurasian haplogroup C in two representatives of the Yamna culture in one of the studied kurgans. Our results indicate the persistence of Mesolithic hunter-gatherer mtDNA lineages in western NPR through the EBA, as well as suggesting a mtDNA lineage continuum connecting the western NPR inhabitants of the Early Metal Ages to the North Pontic Neolithic population groups. Journal of Human Genetics (2017), 1-9 |
|
|
|
Post by Admin on Oct 7, 2018 18:23:23 GMT
 Genetic data suggest that modern European ancestry represents a mosaic of ancestral contributions from multiple waves of prehistoric migration events. Recent studies of genomic variation in prehistoric human remains have demonstrated that two mass migration events are particularly important to understanding European prehistory: the Neolithic spread of agriculture from Anatolia starting ∼9,000 y ago and migration from the Pontic-Caspian Steppe ∼5,000 y ago (1⇓⇓⇓⇓⇓–7). These migrations are coincident with large social, cultural, and linguistic changes, and each has been inferred to have replaced more than half of the contemporaneous gene pool of resident central Europeans. During such events, males and females often experience different demographic histories owing to cultural factors, such as norms regarding inheritance and the residence locations of families in relation to parental residence, social hierarchy, sex-biased admixture, and inbreeding avoidance (8⇓⇓⇓–12). Empirical evidence suggests that sex-specific differences in migration and admixture have shaped patterns of human genomic variation worldwide, with notable examples occurring in Africa, Austronesia, Central Asia, and the Americas (13⇓⇓–16). These sex-specific behaviors leave signatures in the patterns of variation in genetic material that is differentially inherited between males and females in a population. Therefore, contrasting patterns of genetic variation for differentially inherited genetic material can be informative about past sociocultural and demographic events (8⇓⇓⇓–12, 17). Analyses of the maternally inherited mitochondrial DNA (mtDNA) and the paternally inherited Y chromosome have lent differential support to the hypothesis that the Neolithic spread of agriculture from Anatolia occurred through a large population migration rather than a spread of technology (18⇓⇓⇓–22). In general, studies of Y-chromosomal data more than mtDNA have supported Anatolian migration. This pattern of results has been interpreted as evidence for male-biased migration of the population that introduced farming (18, 20, 21). The hypothesis of male-biased migration of farming populations is consistent with ethnographic studies showing a higher frequency of patrilocality in farming than in hunter-gatherer (HG) populations, because an inheritance model through the paternal lineage would favor the persistence of farming-associated Y chromosomes as the source population would have greater flexibility in female mates. Isotopic studies from Neolithic European archeological sites suggest more female than male migration on a local scale, supporting the shift to patrilocality in the region (10, 23). However, genetic evidence has been mixed; both Near Eastern-related mitochondrial and Y-chromosome haplotypes have been observed in European populations, which could indicate comparable male and female migration during the Neolithic spread of agriculture. For example, Haak et al. (22) find that mitochondrial haplotype N1a, associated with Near Eastern farmers, occurs at about ∼25% frequency in Neolithic central Europeans. Later migrations from the steppe, which were previously not accounted for, may have obscured signal and interpretation (22).  Based on archeological data, as well as ancient and modern Y chromosome data, the later migration from the Pontic-Caspian Steppe has also been hypothesized to be male-biased (5, 24⇓⇓⇓⇓–29). In particular, multiple large-scale studies of modern Y-chromosome data infer a rapid growth of R1a and R1b haplotypes ∼5,000 y ago (27⇓–29). Similarly, Haak et al. (5) provide evidence that R1a and R1b were rare in central Europe before ∼4,500 y ago, but common soon thereafter. They also observe multiple R1b haplotypes in ancient Yamnaya individuals from the steppe. Populations in the Pontic-Caspian Steppe region, such as the Yamnaya or Pit Grave culture, are thought to have strong male-biased hierarchy, as inferred by overrepresentation of male burials, male deities, and kinship terms (26, 30). The region is a putative origin for the domesticated horse in Europe, and the culture is known for its use of horse-driven wagons, a potential male-biased mechanism of dispersal into central Europe (30). Recent analytical advances in the understanding of admixture on the autosomes and the sex-specifically–inherited X chromosome and technological advances that have generated genome-wide data from many ancient samples now make it possible to consider the contrasting male and female genetic histories of prehistoric Europe. We test the hypotheses that migrations from Anatolia during the Neolithic transition and from the Pontic Steppe during the late Neolithic/Bronze Age period were male biased.  Fig. 1. Schematic of the admixture history of central European farmers during the Neolithic and Bronze Age. First, a migration from Anatolia occurred during the Neolithic transition, and, second, a late Neolithic/Bronze Age migration occurred from the Pontic-Caspian Steppe to central Europe. In both cases, the migrating population mixed with the contemporaneous local population upon entering central Europe. Fig. 1 provides a schematic of the population admixture events that have previously been inferred (1⇓⇓⇓⇓⇓–7). Previous studies have inferred the relationship between the various ancient populations shown in the figure, but they did not consider a population history model. We compare genetic differentiation of the autosomes and the X chromosome between the migrating and admixed populations for each migration event: Anatolian farmers (AF) to early Neolithic central Europeans (CE) and Pontic Steppe pastoralists (SP) to late Neolithic and Bronze Age central Europeans (BA). We compute the statistic Q(31, 32), which is an estimator of the ratio of effective population size of the X chromosome with the ratio of effective population size of the autosomes based on the FST" role="presentation" style="-webkit-font-smoothing: antialiased; box-sizing: border-box; display: inline; line-height: normal; word-spacing: normal; overflow-wrap: normal; white-space: nowrap; float: none; direction: ltr; max-width: none; max-height: none; min-width: 0px; min-height: 0px; border: 0px; padding: 0px; margin: 0px; position: relative;">FSTFST measure of genetic differentiation (Materials and Methods). Under a simple demographic model with equal male and female effective sizes, Q is expected to be 3/4, because there are three X chromosomes for every four autosomes in the population. Deviations from 3/4 may therefore show sex-biased effective population sizes, which indicate different population histories for males and females. Comparing AF and CE populations for the Neolithic transition, the ratio of X and autosomal differentiation is similar to what is expected for a non–sex-biased process (Q = 0.700; Table 1). In contrast, there is high relative differentiation on the X chromosome between SP and BA populations (Q = 0.237; Table 1), indicating strong male bias during the Pontic Steppe migration. |
|

