|
|
Post by Admin on Mar 5, 2019 17:46:47 GMT
 Fig. 1. Map of the Eurasian steppes. (A) Distribution of the Western (brown) and Eastern (green) steppes and the locations of ancient (red) and modern (black) populations discussed in the text. Population codes are provided in the Dataset S1. A box indicates the location of the LBA burial mounds surveyed in the Arbulag soum of Khövsgöl aimag. (B) Enhanced view of LBA burial mounds (white circles) and burial clusters selected for excavation (boxes a–f) with the number of analyzed individuals in parentheses (SI Appendix, Table S1). (C) Photograph of burial 2009-52 containing the remains of ARS026, a genetic outlier with Western steppe ancestry. Archaeogenetic studies provide evidence that the Eurasian Eneolithic–Bronze Age transition was associated with major genetic turnovers by migrations of peoples from the Pontic-Caspian steppe both in Europe and in central Asia (1⇓⇓⇓–5). The migration of these Western steppe herders (WSH), with the Yamnaya horizon (ca. 3300–2700 BCE) as their earliest representative, contributed not only to the European Corded Ware culture (ca. 2500–2200 BCE) but also to steppe cultures located between the Caspian Sea and the Altai-Sayan mountain region, such as the Afanasievo (ca. 3300–2500 BCE) and later Sintashta (2100–1800 BCE) and Andronovo (1800–1300 BCE) cultures. Although burials typologically linked to the Afanasievo culture have been occasionally reported in Mongolia (6), the genetic profile of Eastern steppe populations, as well as the timing and nature of WSH population expansion and the rise of dairy pastoralism in Mongolia, remain unclear. 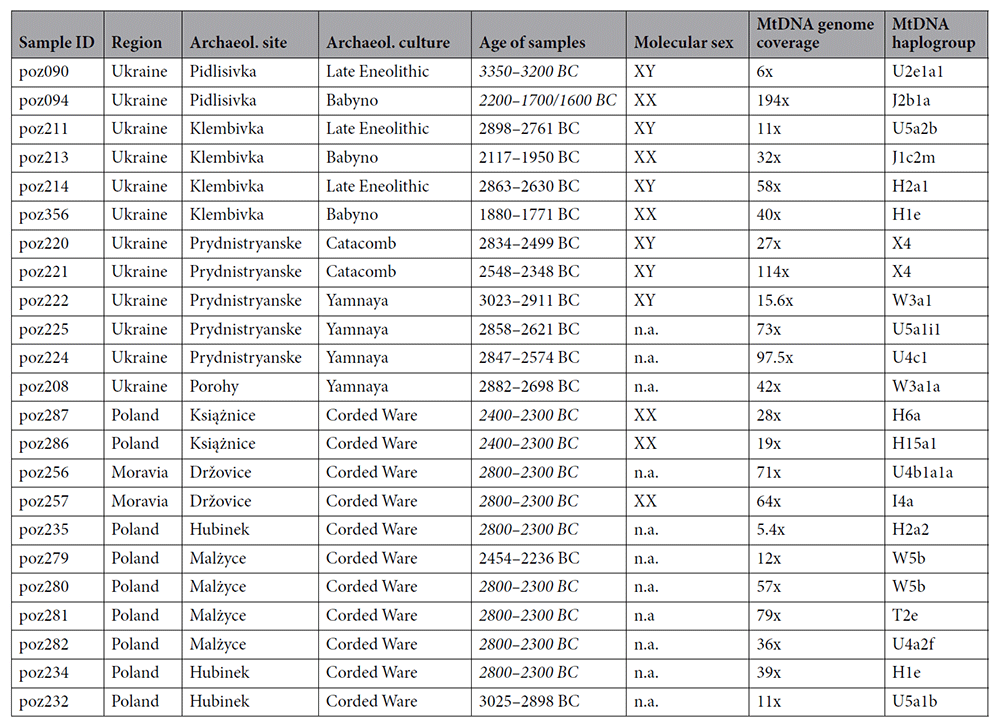 The remarkable demographic success of WSH populations has been linked to mobile pastoralism with dairying (7), a system that efficiently converts cellulose-rich wild grasses into protein- and fat-rich dairy products. Dairy foods provide a rich source of nutrients and fresh water, and function as an adaptive subsistence strategy in cold, dry steppe environments where most crop cultivation is highly challenging. Dairy pastoralism became widely practiced in the eastern Eurasian steppe, the homeland from which subsequent historical nomadic dairying empires, such as the Xiongnu (ca. 200 BCE to 100 CE) and the Mongols (ca. 1200–1400 CE) expanded; however, it is not fully understood when, where, and how this subsistence strategy developed. At Botai, in central Kazakhstan, evidence for Eneolithic dairying has been reported through the presence of ruminant and equine dairy lipids in ceramic residues as early as 3500 BCE (8, 9). In the Altai and Tarim basin, where WSH populations have left strong genetic footprints (1, 3, 10, 11), archaeological evidence supports the presence of dairy products by the early second Millennium BCE and later (8, 12, 13). In the Eastern steppe, however, no direct observations of dairy consumption have been made for a comparable time period, despite the fact that skeletal remains of domestic livestock (such as sheep, goats, cattle, and horses) have been found at Mongolian ritual sites and in midden contexts as early as the 14th century BCE (14⇓⇓–17). In the absence of direct evidence for Bronze Age milk production or consumption on the Eastern steppe, it remains unclear whether these animals are merely ritual in nature or signify a major shift in dietary ecology toward dairy pastoralism, and whether their appearance is connected to possible WSH migrations onto the Eastern steppe. 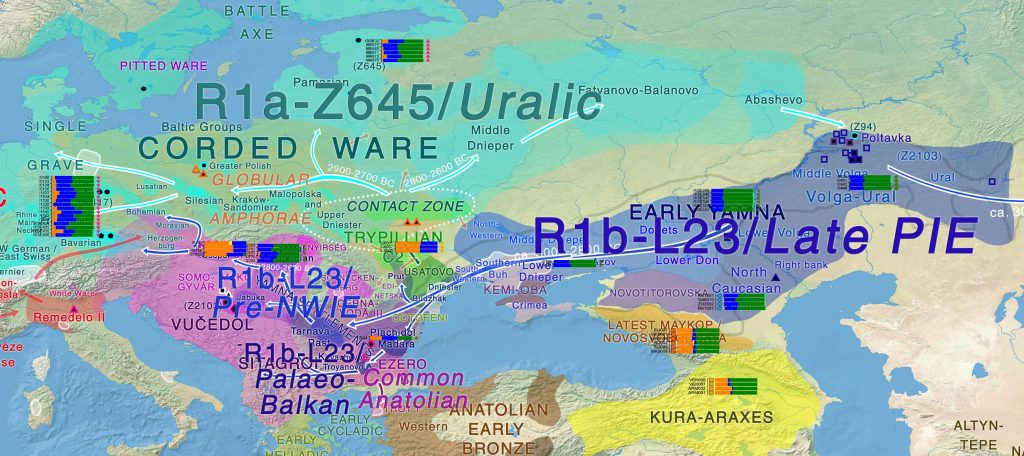 To understand the population history and context of dairy pastoralism in the eastern Eurasian steppe, we applied genomic and proteomic analyses to individuals buried in Late Bronze Age (LBA) burial mounds associated with the Deer Stone-Khirigsuur Complex (DSKC) in northern Mongolia (SI Appendix, Figs. S1–S3 and Table S1). To date, DSKC sites contain the clearest and most direct evidence for animal pastoralism in the Eastern steppe before ca. 1200 BCE (18). Focusing on six distinct burial clusters in Arbulag soum, Khövsgöl aimag, Mongolia (Fig. 1 and SI Appendix, Figs. S1–S3), we produced genome-wide sequencing data targeting ∼1.2M single nucleotide polymorphisms (SNPs) for 22 DSKC-associated individuals directly dated to ca. 1380–975 calibrated BCE (SI Appendix, Fig. S4 and Table S2), as well as sequenced whole genomes for two individuals (>3× coverage). Nine of the individuals in this group yielded sufficient dental calculus for proteomic analysis, and we tested these deposits for the presence of milk proteins using liquid chromatography-tandem mass spectrometry (LC-MS/MS). Overall, our results find that DSKC subsistence strategy included dairying of Western domesticated ruminants, but that there was minimal gene flow between analyzed DSKC populations and WSH groups during the LBA. Thus, in contrast to patterns observed in western Europe where, for example, the arrival of WSH is associated with population replacement and continental-level genetic turnover (5), contact between WSH and Eastern steppe populations is characterized by transcultural transmission of dairy pastoralism in the near absence of demic diffusion. 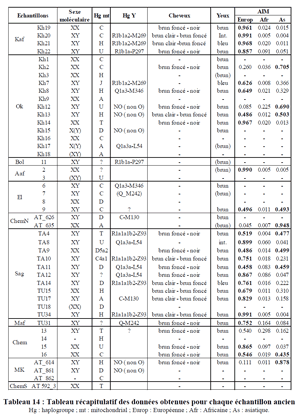 A French study by Clémence Hollard (2014) found that three Afanasievo individuals belonged to R1b, with two classified as R1b-M269. The present work has focused on the settlement of the South Siberia and the Altai mountains during the Bronze Age (III-I millennium BC), period during which the Eurasian Steppes knew a lot of population movements. The molecular analyses were performed on 69 ancient samples. The used strategy consisted in the study of maternal and paternal lineages and autosomal markers informative of biogeographical ancestry (AIM) and physical appearance. Taken together, these results show an evolution of the genetic pool in this area during the Bronze Age. The low effective observed in each cultural group and the funeral recruitment could of course have induced a bias in some analyses. Nevertheless, these molecular data gave new elements to understand the settlement of this region which seems to be a complex process that it will be necessary to deepen with new paleogenetic even paleogenomic studies. |
|
|
|
Post by Admin on Mar 6, 2019 18:26:36 GMT
 Ancient DNA Sequencing and Quality Assessment. We built and sequenced uracil-DNA-glycosylase–half (19), double-indexed Illumina libraries for genomic DNA extracted from teeth or femora from DSKC-associated burials in Khövsgöl, Mongolia. Twenty of 22 libraries exhibited good human DNA preservation, with a mean host endogenous content of 14.9% (range 0.2–70.0%); two libraries yielded very little human DNA (<0.05%) and were excluded from further analysis (SI Appendix, Table S2). Libraries were then enriched for 1.2 million variable sites in the human genome (1240K) using in-solution hybridization (2, 3). All individuals (12 males, 8 females) showed characteristic patterns of chemical modifications typical of ancient DNA (SI Appendix, Fig. S5), and 18 individuals yielded both low estimates of modern DNA contamination (≤1% mitochondrial and nuclear contamination) and sufficient genome coverage for subsequent analysis (0.11× to 4.87× mean coverage for target sites) (SI Appendix, Table S3). No close relative pairs were identified among the ancient individuals (SI Appendix, Fig. S6). Two individuals with high endogenous content on screening (ARS008, 70.0%; ARS026, 47.6%) were deeply sequenced to obtain whole genomes (∼3.3× coverage) (SI Appendix, Table S3). We intersected our ancient data with a published world-wide set of ancient and contemporary individuals (Dataset S1) whose genotypes are determined for 593,124 autosomal SNPs on the Affymetrix HumanOrigins 1 array (20). Characterization of the Genetic Profile of the Khövsgöl Gene Pool. To characterize the genetic profile of DSKC-associated LBA Khövsgöl individuals (Khövsgöls), we performed principal component analysis (PCA) of Eurasian populations (SI Appendix, Fig. S7). PC1 separates eastern and western Eurasian populations, with central and north Eurasian populations falling in an intermediate position (SI Appendix, Fig. S7). PC2 separates eastern Eurasian populations along a north–south cline, with northern Siberian Nganasans and the Ami and Atayal from Taiwan forming the northern and southern end points, respectively. Most LBA Khövsgöls are projected on top of modern Tuvinians or Altaians, who reside in neighboring regions. In comparison with other ancient individuals, they are also close to but slightly displaced from temporally earlier Neolithic and Early Bronze Age (EBA) populations from the Shamanka II cemetry (Shamanka_EN and Shamanka_EBA, respectively) from the Lake Baikal region (SI Appendix, Fig. S7) (4, 21). However, when Native Americans are added to PC calculation, we observe that LBA Khövsgöls are displaced from modern neighbors toward Native Americans along PC2, occupying a space not overlapping with any contemporary population (Fig. 2A and SI Appendix, Fig. S8). Such an upward shift on PC2 is also observed in the ancient Baikal populations from the Neolithic to EBA and in the Bronze Age individuals from the Altai associated with Okunevo and Karasuk cultures (1). These observations are consistent with LBA Khövsgöls and other ancient Siberians sharing more ancestry with Native American-related gene pools than modern populations in the region do.  Fig. 2. The genetic profile of LBA Khövsgöl individuals summarized by PCA and ADMIXTURE. (A) Khövsgöl (Kvs, ARS017, and ARS026) and other ancient individuals (colored symbols) are projected onto the top PCs of modern Eurasian and Native American individuals. Contemporary individuals are marked by gray circles. Mean coordinates for each of the contemporary populations are marked by three-letter codes and by colors assigned to the associated geographic regions. Population codes are provided in Dataset S1 and SI Appendix, Fig. S8. (B) ADMIXTURE results for Khövsgöl and other ancient individuals with K values 9 and 17. In K = 17, the Khövsgöls main cluster is mainly modeled as a mixture of components most enriched in modern northeast Asians (e.g., Nivh) and ancient Siberians (e.g., AG3, Botai, and Okunevo). Notably, two individuals fall on the PC space markedly separated from the others: ARS017 is placed close to ancient and modern northeast Asians, such as early Neolithic individuals from the Devil’s Gate archaeological site (22) and present-day Nivhs from the Russian far east, while ARS026 falls midway between the main cluster and western Eurasians (Fig. 2A). Genetic clustering with ADMIXTURE (23) further supports these patterns (Fig. 2B and SI Appendix, Fig. S9). We quantified the genetic heterogeneity between Khövsgöl individuals by calculating f4 symmetry statistics (24) in the form of f4(chimpanzee, outgroup; Khövsgöl1, Khövsgöl2) for all pairs against 18 outgroups representative of world-wide ancestries (SI Appendix, Fig. S10). As expected, the two outliers did not form a clade with the rest of individuals and therefore we treated each individual separately in subsequent analyses. For the remaining 16 individuals, 14 were merged into a single main cluster based on their minimal genetic heterogeneity. The other two individuals (ARS009 and ARS015) were excluded from this cluster because they broke symmetry with four and two individuals (maximum |Z| = 3.9 and 4.7 SE), respectively, and were also slightly displaced from the others in our PCA (Fig. 2A).  Fig. 3. The genetic affinity of the Khövsgöl clusters measured by outgroup-f3 and -f4 statistics. (A) The top 20 populations sharing the highest amount of genetic drift with the Khövsgöl main cluster measured by f3(Mbuti; Khövsgöl, X). (B) The top 15 populations with the most extra affinity with each of the three Khövsgöl clusters in contrast to Tuvinian (for the main cluster) or to the main cluster (for the two outliers), measured by f4(Mbuti, X; Tuvinian/Khövsgöl, Khövsgöl/ARS017/ARS026). Ancient and contemporary groups are marked by squares and circles, respectively. Darker shades represent a larger f4 statistic. Population codes are provided in Dataset S1; see also SI Appendix, Figs. S11–S14 for further details. Next, we quantified the genetic affinity between our Khövsgöl clusters and world-wide populations by calculating outgroup-f3 statistics with Central African Mbuti as an outgroup (25). For the main cluster, top signals were observed with earlier ancient populations from the Baikal region, such as the early Neolithic and EBA individuals from the Shamanka II cemetry (4), followed by present-day Siberian and northeast Asian populations, such as Negidals from the Amur River basin and Nganasans from the Taimyr peninsula (Fig. 3A and SI Appendix, Fig. S11 A and B). As expected based on their nonoverlapping positions on PCA, however, Khövsgöls do not form a cluster with these high-affinity groups, as shown by f4 symmetry tests in the form of f4(Mbuti, X; Siberian, Khövsgöls). Interestingly, Upper Paleolithic Siberians from nearby Afontova Gora and Mal’ta archaeological sites (AG3 and MA-1, respectively) (25, 26) have the highest extra affinity with the main cluster compared with other groups, including the eastern outlier ARS017, the early Neolithic Shamanka_EN, and present-day Nganasans and Tuvinians (Z > 6.7 SE for AG3) (red shades in Fig. 3B and SI Appendix, Fig. S11 C and D). This extra affinity with so-called “Ancient North Eurasian” (ANE) ancestry (27) may explain their attraction toward Native Americans in PCA, because Native Americans are known to have high proportion of ANE ancestry (20, 25). Main-cluster Khövsgöl individuals mostly belong to Siberian mitochondrial (A, B, C, D, and G) and Y (all Q1a but one N1c1a) haplogroups (SI Appendix, Table S4). |
|
|
|
Post by Admin on Mar 7, 2019 18:16:14 GMT
 Source of ANE Ancestry in the LBA Khövsgöl Population. Previous studies show a close genetic relationship between WSH populations and ANE ancestry, as Yamnaya and Afanasievo are modeled as a roughly equal mixture of early Holocene Iranian/Caucasus ancestry (IRC) and Mesolithic Eastern European hunter-gatherers, the latter of which derive a large fraction of their ancestry from ANE (20, 28). It is therefore important to pinpoint the source of ANE-related ancestry in the Khövsgöl gene pool: that is, whether it derives from a pre-Bronze Age ANE population (such as the one represented by AG3) or from a Bronze Age WSH population that has both ANE and IRC ancestry. To test these competing hypotheses, we systematically compared various admixture models of the main cluster using the qpAdm program (20). Ancient Baikal populations were chosen as a proxy based on both their spatiotemporal and genetic similarities with the Khövsgöl main cluster (Figs. 2 and 3). When the early Neolithic Shamanka_EN is used as a proxy, we find that Baikal+ANE provides a better fit to the main cluster than Baikal+WSH, although no two-way admixture model provides a sufficient fit (P ≥ 0.05) (SI Appendix, Table S5). Adding a WSH population as the third source results in a sufficient three-way mixture model of Baikal+ANE+WSH with a small WSH contribution to the main cluster (e.g., P = 0.180 for Shamanka_EN+AG3+Sintashta with 3.7 ± 2.0% contribution from Sintashta) (Fig. 4 and SI Appendix, Table S6).  Fig. 4. Admixture modeling of Altai populations and the Khövsgöl main cluster using qpAdm. For the archaeological populations, (A) Shamanka_EBA and (B and C) Khövsgöl, each colored block represents the proportion of ancestry derived from a corresponding ancestry source in the legend. Error bars show 1 SE. (A) Shamanka_EBA is modeled as a mixture of Shamanka_EN and AG3. The Khövsgöl main cluster is modeled as (B) a two-way admixture of Shamanka_EBA+Sintashta and (C) a three-way admixture Shamanka_EN+AG3+Sintashta. Details of the admixture models are provided in SI Appendix, Tables S5 and S6. Using the temporally intermediate EBA population Shamanka_EBA, we can narrow down the time for the introduction of WSH ancestry into the main cluster. Shamanka_EBA is modeled well as a two-way mixture of Shamanka_EN and ANE (P = 0.158 for Shamanka_EN+AG3) (Fig. 4) but not as a mixture of Shamanka_EN and WSH (P ≤ 2.91 × 10−4) (SI Appendix, Table S5), suggesting no detectable WSH contribution through the early Bronze Age. Similar results are obtained for other Late Neolithic and EBA populations from the Baikal region (SI Appendix, Table S5). In contrast, the Khövsgöl main cluster is modeled well by Shamanka_EBA+WSH but not by Shamanka_EBA+ANE (P ≥ 0.073 and P ≤ 0.038, respectively) (SI Appendix, Table S5). A three-way model of Shamanka_EBA+ANE+WSH confirms this by providing the ANE contribution around zero (SI Appendix, Table S6). The amount of WSH contribution remains small (e.g., 6.4 ± 1.0% from Sintashta) (Fig. 4 and SI Appendix, Table S5). Assuming that the early Neolithic populations of the Khövsgöl region resembled those of the nearby Baikal region, we conclude that the Khövsgöl main cluster obtained ∼11% of their ancestry from an ANE source during the Neolithic period and a much smaller contribution of WSH ancestry (4–7%) beginning in the early Bronze Age. Admixture Testing of Genetic Outliers. Using the same approach, we obtained reasonable admixture models for the two outliers, ARS017 and ARS026. The eastern outlier ARS017, a female, shows an extra affinity with early Neolithic individuals from the Russian far east (Devil’s Gate) (22) and in general with contemporary East Asians (e.g., Han Chinese) compared with the Khövsgöl main cluster (Fig. 3B and SI Appendix, Fig. S12). ARS017 is also similar to Shamanka_EN in showing no significant difference in qpAdm (SI Appendix, Fig. S12 and Table S7). Using contemporary East Asian proxies, ARS017 is modeled as a mixture of predominantly Ulchi and a minor component (6.1–9.4%) that fits most ancient western Eurasian groups (P = 0.064–0.863) (SI Appendix, Table S7). This minor Western component may result from ANE ancestry; however, given the minimal western Eurasian contribution, we do not have sufficient power to accurately characterize this individual’s western Eurasian ancestry. The Western outlier ARS026, a male dating to the end of the radiocarbon series, has the highest outgroup-f3 with the main LBA Khövsgöl cluster, with extra affinity toward Middle Bronze Age (MBA) individuals from the Sintashta culture (Fig. 3B and SI Appendix, Fig. S13) (1). DNA recovered from this individual exhibited expected aDNA damage patterns (SI Appendix, Fig. S5) but was otherwise excellently preserved with >47% endogenous content and very low estimated contamination (1% mitochondrial; 0.01% nuclear). ARS026 is well modeled as a two-way mixture of Shamanka_EBA and Sintashta (P = 0.307; 48.6 ± 2.0% from Sintashta) (SI Appendix, Table S7). Similar to ARS026, contemporaneous LBA Karasuk individuals from the Altai (1400–900 BCE) (1, 29) also exhibit a strong extra genetic affinity with individuals associated with the earlier Sintashta and Andronovo cultures (SI Appendix, Fig. S14). Although two-way admixture models do not fit (P ≤ 0.045) (SI Appendix, Table S8), the Karasuk can be modeled as a three-way mixture of Shamanka_EBA/Khövsgöl and AG3 and Sintashta, suggesting an eastern Eurasian source with slightly higher ANE ancestry than those used in our modeling (P ≥ 0.186) (SI Appendix, Table S8). Like ARS026, admixture coefficients for the Karasuk suggest that MBA/LBA groups like the Sintashta or Srubnaya are a more likely source of their WSH ancestry than the EBA groups, like the Yamnaya or Afanasievo. Notably, Karasuk individuals are extremely heterogeneous in their genetic composition, with the genetically easternmost Eurasian individual nearly overlapping with the EBA Baikal groups (Fig. 2A and SI Appendix, Figs. S7 and S8). Earlier groups, such as the Afanasievo, Sintashta, and Andronovo, are mostly derived from WSH ancestries, and this may suggest that admixture in the Altai-Sayan region only began during the LBA following a long separation since the Eneolithic. Although ARS026 exhibits substantial WSH ancestry, strontium isotopic values obtained from his M3 enamel resemble local fauna and fall within the range of the main Khövsgöl cluster (SI Appendix, Fig. S15 and Table S9); however, because the enamel this individual also exhibited elevated manganese levels, postmortem trace element alteration from soil could not be excluded. |
|
|
|
Post by Admin on Mar 8, 2019 18:26:43 GMT
Dairy Subsistence and Lactase Persistence. Contemporary Mongolia has a dairy- and meat-based subsistence economy, and to more precisely understand the role of dairy products in the diets of present-day mobile pastoralists in Khövsgöl aimag, we conducted a detailed nutritional investigation of summer and winter diets. We find that dairy-based foods contribute a mean of 35% total dietary energy, 36–40% total carbohydrate, 24–31% total protein, and 39–40% total fat to rural summer diets in Khövsgöl aimag, with liquid milk and dairy product consumption of 216–283 and 172–198 g/d, respectively (SI Appendix, Table S10 and Dataset S2). Despite the importance of dairying today, its origins in Mongolia are poorly understood. Given the limited WSH ancestry of the main Khövsgöl cluster, we sought to determine if dairy pastoralism was practiced by this putatively pastoralist LBA population by testing for the presence of milk proteins (30) in the dental calculus of these individuals. We extracted proteins from 12 dental calculus samples representing 9 individuals (SI Appendix, Table S11) and analyzed tryptic peptides using LC-MS/MS (31). Observed modifications included deamidation (N, Q) and oxidation (P, M) (SI Appendix, Table S12). All protein identifications were supported by a minimum of two peptides across the dataset, and only peptides with an E value ≤ 0.001 were assigned; the estimated peptide false-discovery rate (FDR) across the full dataset was 1.0%, and protein FDR was 4.6%. Milk proteins were detected in seven of the nine individuals analyzed (SI Appendix, Table S13 and Dataset S3), confirming that dairy foods were consumed as early as 1456 BCE (1606–1298 BCE, 95% probability of the earliest directly dated individual) (SI Appendix, Fig. S4 and Table S2). Specifically, we detected the milk whey protein β-lactoglobulin (Fig. 5 A and B) and the curd protein α-S1-casein, with peptides matching specifically to sheep (Ovis), goat (Capra), Caprinae, Bovinae, and a subset of Bovidae (Ovis or Bovinae) (Fig. 5C, SI Appendix, Table S13, and Dataset S3). These peptides exhibited asparagine and glutamine deamidation, as expected for ancient proteins (32), and the frequency and distribution of recovered β-lactoglobulin (Fig. 5B) and α-S1-casein peptides closely matched that empirically observed for modern bovine milk (33), thereby providing additional protein identification support through appropriate proteotypic behavior.  Fig. 5. Presence of ruminant β-lactoglobulin and α-S1-casein milk protein in LBA Khövsgöl dental calculus. (A) B- and Y-ion series for one of the most frequently observed β-lactoglobulin peptides, TPEVD(D/N/K)EALEKFDK, which contains a genus-specific polymorphic residue: D, Bos; N, Ovis; K, Capra. See SI Appendix, Fig. S16 for peptide and fragment ion error distribution graphs. (B) Alignment of observed peptides to the 178 amino acid β-lactoglobulin protein, with peptide taxonomic source indicated by color. Trypsin cut sites are indicated by gray ticks. The position and empirically determined observation frequency of BLG peptides for bovine milk are shown as a heatmap scaled from least observed peptides (light gray) to most frequently observed peptides (dark red), as reported in the Bovine PeptideAtlas (34). Inset displays a 3D model of the β-lactoglobulin protein with observed peptide positions highlighted in black. (C) Taxonomically assigned β-lactoglobulin (black) and α-S1-casein (gray) peptides presented as scaled pie charts on a cladogram of Mongolian dairy domesticates. Bracketed numbers represent the number of peptides assigned to each node. Ruminant milk proteins were well supported, but no cervid, camelid, or equid milk proteins were identified. Given the evidence for dairy consumption by the LBA Khövsgöl population, we sought to determine if the dairy-adaptive -13910*T (rs4988235) lactase persistence (LP) allele found today in Western steppe (34) and European (35) populations was present among LBA Khövsgöls dairy herders, and we examined this position in our SNP-enriched dataset. The -13910*T LP allele was not found in the LBA Khövsgöls (SI Appendix, Fig. S17 and Table S14), and additionally all observed flanking sequences in the lactase transcriptional enhancer region contained only ancestral alleles. Despite the limited gene flow between the Western and Eastern steppes, dairy pastoralism was nevertheless adopted by local non-WSH populations on the Eastern steppe and established as a subsistence strategy by 1300 BCE. Ruminant milk proteins were identified in the dental calculus of most of the tested LBA Khövsgöl individuals, and all identified milk proteins originated from ruminants, specifically the Western dairy domesticates sheep, goat, and Bovinae. These findings suggest that neighboring WSH populations directly or indirectly introduced dairy pastoralism to local indigenous populations through a process of cultural exchange. Further research on other regional cultures in Mongolia, such as Chemurchek, Hemsteg, and Ulaanzuukh, is needed to determine if this pattern of cultural adoption observed among DSKC sites is broadly shared across other Bronze Age cultures throughout the Eastern steppe. 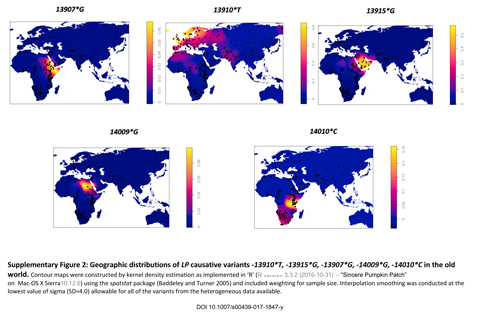 Bronze Age trade and cultural exchange are difficult to observe on the Eastern steppe, where mobile lifestyles and ephemeral habitation sites combine to make household archaeology highly challenging. Burial mounds are typically the most conspicuous features on the landscape, and thus much of Mongolian archaeology is dominated by mortuary archaeology. However, unlike WSH, whose kurgans typically contain a range of grave goods, many LBA mortuary traditions on the Eastern steppe did not include grave goods of any kind other than ritually deposited animal bones from horse, deer, and bovids. Given that Mongolian archaeological collections are typically dominated by human remains with limited occupational materials, the ability to reconstruct technological exchange, human–animal interaction, and secondary product utilization through the analysis of proteins preserved in dental calculus represents an important advance. The 3,000-y legacy of dairy pastoralism in Mongolia poses challenging questions to grand narratives of human adaptation and natural selection (36). For example, despite evidence of being under strong natural selection (36), LP was not detected among LBA Khövsgöls, and it remains rare (<5%) in contemporary Mongolia even though levels of fresh and fermented dairy product consumption are high (35). Recent studies in Europe and the Near East have found that dairying preceded LP in these regions by at least 5,000 y, suggesting that LP may be irrelevant to the origins and early history of dairying (36). As a non-LP dairying society with a rich prehistory, Mongolia can serve as a model for understanding how other adaptations, such as cultural practices or microbiome alterations (37), may be involved in enabling the adoption and long-term maintenance of a dairy-based subsistence economy. Early herding groups in Mongolia present a historical counter-example to Europe in which WSH migrations resulted in cultural exchange rather than population replacement, and dairying was maintained for millennia without the introgression or selection of LP alleles.NAS November 27, 2018 115 (48) E11248-E11255 www.pnas.org/content/pnas/early/2018/10/31/1813608115.full.pdf |
|
|
|
Post by Admin on Mar 12, 2019 18:43:49 GMT
 The origin of the Yamnaya The Yamnaya people originated on the Caspian steppes where they lived as pastoralists and herders, using wagons as mobile homes. From burial pits archaeologists have found extensive use of thick plant mats and felt covers. Their economy was based on meat, dairy products and fish, they were tall and rather healthy with little caries in their teeth. No agriculture is documented. Barrows were aligned in groups forming lines in the landscape to mark seasonal routes and after death diseased people were put into individual graves under small family barrows. Their burial ritual thus embodied a new perception of the individual and of small monogamous family groups as the foundation of society. The continent encountered by the Yamnaya people around 3000 BC had seen a decline in the agrarian Stone Age societies, thereby allowing space for incoming migrants. This decline was probably the result of a widespread plague from Siberia to the Baltic. "The disease dynamic here may have been comparable to the European colonization process in America after Christopher Columbus", says Kristiansen. "Perhaps Yamnaya brought plague to Europe and caused a massive collapse in the population".  Genetic structure of ancient Europe and the Pontic-Caspian steppe "Black Youth" as migrating males In the new synthesis article, Kristiansen and colleagues argue for a dominance of males during the early phase after the migrations, and correspond to the old Indo-European mythology of later times. These sources talk about war-bands of youths – called "Black Youth" – who were employed in pioneer migrations as a dynamic force. Evidence from strontium isotopic analyses, published in 2016 by Kristiansen together with Douglas Price and Karl Goran Sjogren, showed that a majority of the women in Corded Ware burials in south Germany were non-locals who had married in from Neolithic societies, since they had a Neolithic diet in their childhood. These results now form part of the new synthesis. Professor Kristian Kristiansen says: "Existing archaeological evidence of a strong 90% male dominance in the early phase of the Corded Ware/Single Grave Culture settlement in Jutland, Denmark, and elsewhere can now be explained by the old Indo-European tradition of war bands of young males who did not have any inheritance to look forward to. Therefore they were probably more willing to make a career as migrating war bands." These Neolithic women also brought new knowledge of pottery production, and started to imitate pottery containers made of wood from the Yamnaya migrants. In this way a new pottery culture was created called Corded Ware, because of the cord impressions around the neck of the pots. They were made for beer drinking, and the new migrants also learned how to grow barley from the in-married Neolithic women in order to produce beer.  Rapid genetic changeover Eske Willerslev undertook the ancient DNA analyses together with Morten Allentoft and Martin Sikora. Professor Willerslev says: "In our big Bronze Age study, published in 2015, we were astonished to see how strong and fast the genetic changeover was from the Neolithic to the Corded Ware. There was a heavy reduction of Neolithic DNA in temperate Europe, and a dramatic increase of the new Yamnaya genomic component that was only marginally present in Europe prior to 3000 BC. Moreover, the apparent abruptness with which this change occurred indicates that it was a large-scale migration event, rather than a slow periodic inflow of people."  Allele frequencies for putatively positively selected SNPs. New words and new Proto-Germanic dialect The Yamnaya brought the Indo-European languages into Bronze Age Europe, but as herders, they did not have words for crops or cultivation, unlike the Neolithic farmers. As the Corded Ware Culture developed it adopted words related to farming from the indigenous Neolithic people, which they were admixing with. Guus Kroonen, a historical linguist, was able to demonstrate that these new words did not belong to the original Indo-European languages. Therefore it was possible to conclude that the Neolithic people were not speaking an Indo-European language, as did the Yamnaya migrants. Thus, the process of genetic and cultural admixture was accompanied by a process of language admixture, creating the foundations for later Germanic languages, termed Proto-Germanic. The birth of the Bronze Age The Yamnaya migrations from the Pontic-Caspian steppe into temperate Europe changed the course of history: they brought not only a new language, but also new ideas about how society was organized around small monogamous families with individual ownership to animals and land. This new society became the foundation for the Bronze Age, and for the way European societies continued to develop to the present. The paper Re-theorising mobility and the formation of culture and language among the Corded Ware Culture in Europe by Kristiansen, Allentoft, Frei, Iversen, Johannsen, Kroonen, Pospieszny, Price, Rasmussen, Sjögren, Sikora and Willerslev is published in the journal Antiquity 4 April 2017. The Bronze Age of Eurasia (around 3000–1000 BC) was a period of major cultural changes. However, there is debate about whether these changes resulted from the circulation of ideas or from human migrations, potentially also facilitating the spread of languages and certain phenotypic traits. We investigated this by using new, improved methods to sequence low-coverage genomes from 101 ancient humans from across Eurasia. We show that the Bronze Age was a highly dynamic period involving large-scale population migrations and replacements, responsible for shaping major parts of present-day demographic structure in both Europe and Asia. Our findings are consistent with the hypothesized spread of Indo-European languages during the Early Bronze Age. We also demonstrate that light skin pigmentation in Europeans was already present at high frequency in the Bronze Age, but not lactose tolerance, indicating a more recent onset of positive selection on lactose tolerance than previously thought. Nature volume 522, pages 167–172 (11 June 2015) |
|

