|
|
Post by Admin on Mar 14, 2019 19:31:23 GMT
 The Pontic-Caspian steppe (PCS), stretching from the southern Urals to the western North Pontic lands, was the stage of various demographic changes in the past, and several of those remain unknown. During the Bronze and Iron Age, the area was inhabited by a succession of nomadic populations that had significant impact on the cultural development of both Asia and Europe (1, 2). Possibly the best known of these groups is the Yamnaya. Recent genomic studies have revealed cross-continental Early Bronze Age migrations (~3000 BCE) of the nomadic people associated with the Yamnaya horizon (3, 4). The migration introduced the Caucasus genetic component to the genetic landscape of Europe. In Central Europe, Yamnaya ancestry first appeared among people from the Corded Ware complex and has since been found in many subsequent ancient and present-day populations. However, the Pontic-Caspian steppe was critical not only for Early Bronze Age Yamnaya migrations but also because of succeeding movements and population transformations that took place in the developed classical stage of the Late Bronze and Iron Ages between 1800 BCE and 400 CE. This period covered the development of the Srubnaya and Alakulskaya Cultures (~1800–1200 BCE), associated with small settlement sites distributed from the Urals to the Dnieper valley (1). From around 1000 BCE, pre-Scythian nomadic populations started to appear in the western Pontic-Caspian steppe including the Cimmerians known from historical sources (5). Despite regional variation and local peculiarities, the Cimmerians were not associated with any uniform type of archaeological material culture (6). In the seventh century BCE, they were succeeded by the Scythians, who plausibly pushed the Cimmerians into Asia Minor (7). Between 700 and 300 BCE, the Scythians, representing mobile pastoral nomads of a new militaristic type (1), dominated the Pontic-Kazakh steppe, occupying an area from the Altai to the Carpathian Mountains. Their decline began around 300 BCE and was caused by intensifying hostile relations with the Macedonians in the West and the invasion of the Sarmatians from the East. The Sarmatians and the Scythians are thought to have coexisted for a few centuries, but eventually, the former group prevailed (2), resulting in the Scythian downfall. The Sarmatians are believed to comprise a number of groups of similar nomadic background (8), and they became the politically most influential force within the eastern fringes of the Roman Empire at the time. Their decline (~400 CE) was associated with the attack of the Goths and the subsequent invasion of the Huns (8). The genomic structure of the Bronze and Iron Age (1800 BCE–400 CE) populations in the Pontic-Caspian steppe has not been fully resolved. While earlier genomic studies have suggested close links between the Srubnaya and the central European Late Neolithic and Bronze Age populations (9), our knowledge of the genetic origins of the Cimmerians is limited. Genetic analyses of maternal lineages of Scythians suggest a mixed origin and an east-west admixture gradient across the Eurasian steppe (10–12). The genomics of two early Scythian Aldy-Bel individuals (13) showed genetic affinities to eastern populations of Central Asia (12). However, population interactions and the origin of Scythians of the Pontic-Caspian steppe remain poorly understood. Similarly, little is known about the origins and genetic affinities of the Sarmatians. Genomic studies suggest that the latter group may have been genetically similar to the eastern Yamnaya and Poltavka Bronze Age groups (12). To investigate the demographic dynamics in the Pontic-Caspian steppe, we generated and analyzed genomes of the Late Bronze and Iron Age individuals from the region (Fig. 1, A and B).  Fig. 1 Radiocarbon ages and geographical locations of the ancient samples used in this study. We produced genome-wide sequence data with genome coverage between 0.01× and 2.9× per individual for 35 Bronze Age and Iron Age individuals from the Pontic-Caspian steppe from four chronologically sequential cultural groups, which comprise Srubnaya-Alakulskaya individuals (n = 13), Cimmerians (n = 3), Scythians (n = 14), and Sarmatians (n = 5), with radiocarbon dates between ca. 1900 BCE and 400 CE (Fig. 1, A and B; tables S1 to S3; and fig. S1, A and B). All DNA libraries displayed damage patterns typical of ancient DNA (fig. S2) (14). To ensure data integrity, we calculated mitochondrial DNA (mtDNA)–based contamination levels using distribution of private polymorphisms in mtDNA (15) and a Bayesian likelihood method (16). The former yielded point estimates of contamination between 0 and 10% [95% confidence intervals (CIs) between 0 and 17%], and the latter method revealed that all individuals carried sequences with >89% probability of being authentic (table S4). Thus, we included all sequenced individuals in the downstream analyses. Late Bronze Age (LBA) Srubnaya-Alakulskaya individuals carried mtDNA haplogroups associated with Europeans or West Eurasians (17) including H, J1, K1, T2, U2, U4, and U5 (table S3). In contrast, the Iron Age nomads (Cimmerians, Scythians, and Sarmatians) additionally carried mtDNA haplogroups associated with Central Asia and the Far East (A, C, D, and M) (table S3) (11, 18). The absence of East Asian mitochondrial lineages in the more eastern and older Srubnaya-Alakulskaya population suggests that the appearance of East Asian haplogroups in the steppe populations might be associated with the Iron Age nomads, starting with the Cimmerians. The Y chromosome haplogroup variation in 17 of 18 males was limited to two major haplogroup lineages within the macrohaplogroup “R” (table S3). The Srubnaya-Alakulskaya individuals carried the Y haplogroup R1a, which showed a major expansion during the Bronze Age (19). It has previously been found in Bronze Age individuals from the Krasnoyarsk Kurgan in Siberia (20). The Iron Age nomads mostly carried the R1b Y haplogroup, which is characteristic of the Yamnaya of the Russian steppe (4). An exception was a Cimmerian individual (cim358) who carried the Q1* lineage associated with the east (table S3). Genetic relationships between Eurasian steppe nomads and present-day populations PCA on the autosomal genomic data (Fig. 1C and table S5) revealed the following: (i) Srubnaya-Alakulskaya individuals exhibited genetic affinity to northern and northeastern present-day Europeans (fig. S3), and these results were also consistent with outgroup f3 statistics (table S6 and fig. S4A). (ii) The Cimmerian individuals, representing the time period of transition from Bronze to Iron Age, were not homogeneous regarding their genetic similarities to present-day populations according to the PCA. F3 statistics confirmed the heterogeneity of these individuals in comparison with present-day populations (table S6 and figs. S3 and S4C). (iii) The Scythians reported in this study, from the core Scythian territory in the North Pontic steppe (12), showed high intragroup diversity. In the PCA, they are positioned as four visually distinct groups compared to the gradient of present-day populations (Fig. 1C): (i) A group of three individuals (scy009, scy010, and scy303) showed genetic affinity to north European populations, hereafter referred to as a north European (NE) cluster. (ii) A group of four individuals (scy192, scy197, scy300, and scy305) showed genetic similarities to southern European populations, hereafter referred to as a south European (SE) cluster. (iii) A group of three individuals (scy006, scy011, and scy193) located between the genetic variation of Mordovians and populations of the North Caucasus, hereafter referred to as a steppe cluster (SC). In addition, one Srubnaya-Alakulskaya individual (kzb004), the most recent Cimmerian (cim357), and all Sarmatians fell within this cluster. In contrast to the Scythians, and despite being from opposite ends of the Pontic-Caspian steppe, the five Sarmatians grouped close together in this cluster. (iv) A group of three Scythians (scy301, scy304, and scy311) formed a discrete group between the SC and SE and had genetic affinities to present-day Bulgarian, Greek, Croatian, and Turkish populations, hereafter referred to as a central cluster (CC). All PCA results were consistent with outgroup f3 statistics (table S6 and figs. S3 and S4, B and D). Finally, one individual from a Scythian cultural context (scy332) is positioned outside of the modern West Eurasian genetic variation (Fig. 1C) but shared genetic drift with East Asian populations (table S6 and fig. S4B). Genetic relationships between Srubnaya-Alakulskaya and other ancient populations The Bronze Age in Eurasia was a dynamic period with several human groups participating in different migratory processes. As a result, there were extensive interactions between European Bronze Age populations (3). The Srubnaya-Alakulskaya individuals, originating from a site of cultural dualism in the forest steppe of the Trans-Volga region, were genetically similar to the previously published Srubnaya and Andronovo individuals from the Pontic-Kazakh steppe (3, 9) and to the European Bronze Age groups, including individuals of the Corded Ware, Unetice, and Bell Beaker complexes (Fig. 1C and fig. S5). Consistent with other Bronze Age populations, the Srubnaya-Alakulskaya individuals were positioned between the genetic variation of the European Mesolithic and the Near East Neolithic populations, being closer to the former and especially to the east European hunter-gatherers (Fig. 1C and figs. S6 and S8). These individuals had higher genetic affinity to Scythians compared to other Iron Age groups (fig. S9). To test whether our Bronze Age sample from a cultural mixing zone between the Srubnaya and the Alakulskaya complexes shared more genetic drift with previously published representatives of Srubnaya, we calculated f4 statistics of f4(Yoruba, SrubnayaX, SrubnayaY, BronzeX), where “BronzeX” refers to a Bronze Age Russian population. The test revealed that the Srubnaya-Alakulskaya population formed a clade together with Andronovo, Afanasievo, and Sintashta to the exclusion of the previously published data of other Srubnaya individuals (tables S7 and S8). Furthermore, to trace the plausible origin the Caucasus genetic component identified in Srubnaya-Alakulskaya individuals, we adopt the f4 statistics in the form of f4(Yoruba, Srubnaya-Alakulskaya, PopulationX, Yamnaya), where PopulationX was one of the Eurasian Bronze Age populations. The results showed that Srubnaya-Alakulskaya formed a clade together with Yamnaya to the exclusion of other Bronze Age populations from Russia, Armenia, Jordan, and Hungary. This finding indicates that the Caucasus genetic contribution to the Srubnaya-Alakulskaya individuals was mediated by steppe ancestry instead of originating from the Levant (table S9). Both mean f3 statistics within populations and conditional nucleotide diversity (21) revealed that the genetic diversity was highest in the LBA Srubnaya-Alakulskaya population from the southern Ural region compared to all other Eurasian Bronze Age populations (Fig. 2, A and B, and table S10).  Fig. 2 Genetic diversity and ancestral components of Srubnaya-Alakulskaya population. To evaluate the ancestral components of Srubnaya-Alakulskaya, we conducted ADMIXTURE analysis for K = 2 to K = 15 ancestral clusters (fig. S10A). K = 15 clusters revealed that Srubnaya-Alakulskaya individuals consisted of two major ancestral components; first, an “orange” component predominantly found in west and NE hunter-gatherers (WHG) and in present-day NE populations, and second, a “light green” component typical of Caucasus hunter-gatherers (CHG) found in south Asian (SA) modern populations. The component associated with Neolithic groups and present-day Near Eastern populations (NEN, “dark red”) contributed less to our Srubnaya-Alakulskaya individuals compared to the European Bronze Age populations (Fig. 2C and fig. S10B). |
|
|
|
Post by Admin on Mar 15, 2019 18:43:38 GMT
Genetic relationships between the Iron Age nomads and other ancient populations In the 10th century BCE, during the transition from the Bronze Age to the Iron Age, the Cimmerians appear in the Pontic-Caspian steppe. In the PCA, the chronologically youngest Cimmerian individual (cim357) grouped within the SC including all Sarmatians, one Srubnaya-Alakulskaya individual, and three Scythian individuals. The other Cimmerian individuals were positioned in close proximity to a number of eastern Iron Age individuals from the Altai region (3), as well as Aldy-Bel and Zevakino-Chillikta individuals of the Altai and Western Mongolia regions (Fig. 1C and fig. S9) (12). We tested whether Cimmerians formed a clade with the subsequent Iron Age populations including Scythians and Sarmatians, to the exclusion of Bronze Age populations by calculating f4 statistics in the form of f4(Yoruba, Cimmerians; Srubnaya&Scythians&Sarmatians, BronzeAgePopulation), showing that the Cimmerians shared more drift with the Bronze Age populations of Russia compared to the Srubnaya/-Alakulskaya but not compared to Scythians or Sarmatians (Fig. 3A and tables S11, S12, and S16). The Cimmerians shared more drift with the far eastern Karasuk population compared to the geographically more closely located Srubnaya/-Alakulskaya population (table S16), corroborating the existence of the “Karasuk-Cimmerian cultural-historical community” (22). Pairwise mismatch estimates revealed a slightly higher genetic diversity in Cimmerians compared to that of the Srubnaya-Alakulskaya population (table S10). ADMIXTURE analysis (K = 15) revealed that Cimmerian individuals carried predominantly the WHG and CHG components similar to the Srubnaya individuals, but they also carried other ancestral components including NEN (dark red), Northeast Asian (NEA) (“dark green”), and Southeast Asian (SEA, “light blue”) components. From the oldest to the most recent sample, the amount of NEN component increased, while the NEA and SEA components decreased through time in the Cimmerians (Fig. 3D and fig. S10). The admixture results were further tested with f3 statistics, which were consistent with the observed pattern by means of shared genetic drift between the Cimmerians and the tested populations used as proxies for different genetic components identified in admixture analyses (table S13). These results implicate a more eastern origin of the Cimmerians compared to that of the Srubnaya-Alakulskaya population and an increased amount of NEN-originated admixture in these individuals through time.  Fig. 3 Genetic relationship between Srubnaya-Alakulskaya population and Iron Age nomads. Visual summary of f4 statistics of a form f4(YRI, TestPop, Pop X, Srubnaya), where TestPop is (A) Cimmerian; (B) Scythian and (C) Sarmatian. (D) Admixture selection: Our ancient individuals and Iron Age individuals from closely related populations (K = 15). K = 15 was chosen to display since it shows SA (lilac) and NEA (dark green) components in addition to the other components. Srubnaya-Alakulskaya population is named as Srubnaya in the panels. IA, Iron Age (fig. S10). In the seventh century BCE, the Scythians appeared in the North Pontic region. Iron Age western Scythians displayed slightly higher intragroup diversity compared to that of the Bronze Age groups and formed four discrete clusters including NE, SE, SC, and CC clusters (Fig. 1C and table S10). Scythians belonging to the SE cluster were closer to Hungarian Bronze Age and Iron Age individuals including Vatya and Maros. The NE Scythian cluster fell close to the Iron Age individuals from modern-day Montenegro and Sweden (fig. S9). The SC Scythians further grouped with Early Sarmatians (12) and the Iron Age Scythian from modern-day Hungary (23). It has been hypothesized by Terenozhkin that Scythians reached the Pontic-Caspian steppe region from Central Asia (for example, from Andronovo) (7). To formally test this hypothesis, we calculated f4 statistics in the form of f4(Yoruba, Scythian, Srubnaya, BronzeAgeX), where “BronzeAgeX” refers to either Andronovo, Sintashta, or Afanasievo. This analysis revealed that the western Scythians (tested either as a single population or four different clusters) formed a clade together with Andronovo and two other Andronovo-originated populations to the exclusion of Srubnaya-Alakulskaya from the southern Urals (Fig. 3B and table S14), supporting the Central Asian origin of the western Scythians. Furthermore, the western Scythians shared more drift with Andronovo, Afanasievo, Sintashta, and Mezhovskaya to the exclusion of Yamnaya (table S15). While the eastern Scythians from an earlier study (12) formed a clade together with Srubnaya-Alakulskaya to the exclusion of Yamnaya, the western Scythians of the present study did not show this pattern. Only when compared with Karasuk and Okunevo, Yamnaya seemed closer (although scy332 is an exception), which is in line with a shared steppe origin of the Yamanya and the western nomads (table S15). ADMIXTURE analysis at K = 15 revealed that the Scythians belonging to the NE cluster had a more visible West Eurasian component that is highest in the northern populations, while Scythians that belong to the SE cluster had a more visible NEN genetic component, most commonly found in Neolithic Anatolian individuals. The NEN component was also found in individuals falling into the CC, who additionally carried an SA component (“lilac”) shared with the Cimmerians, Sarmatians, and two more Scythian individuals (scy193 and scy006). Scy332 had a SEA (“light blue”) component and was most similar to the Karasuk individuals (Fig. 3 and fig. S10, A and B). The five Sarmatians grouped close together in the SC (Fig. 1C), but they also had the highest pairwise mismatch estimates compared to other Iron Age nomads, suggesting a larger effective population size (table S10). F4(Yoruba, Sarmatians, PopulationX, Cimmerian), where the PopulationX is Yamnaya, Srubnaya, or Karasuk, revealed that Sarmatians formed a clade together with all these three populations to the exclusion of Cimmerians (table S18). Multiway f4 statistics testing the relationships between Srubnaya/-Alakulskaya, Cimmerians, and Scythians revealed that both Scythians and Cimmerians formed a clade together with Srubnaya/-Alakulskaya to the exclusion of the other population and that Srubnaya/-Alakulskaya was closer to Scythians among the two (Fig. 3C and table S17). The results point to the presence of a deep shared ancestry of all Iron Age nomadic groups associated with Bronze Age populations of the steppe, which, however, is not equivalent with a direct genetic continuity between Srubnaya-Alakulskaya and the western Scythians.  DISCUSSION The origins of the four steppe populations The Bronze Age Srubnaya-Alakulskaya individuals from Kazburun 1/Muradym 8 presented genetic similarities to the previously published Srubnaya individuals. However, in f4 statistics, they shared more drift with representatives of the Andronovo and Afanasievo populations compared to the published Srubnaya individuals. Those apparently West Eurasian people lacked significant Siberian components (NEA and SEA) in ADMIXTURE analyses but carried traces of the SA component that could represent an earlier connection to ancient Bactria. The presence of an SA component (as well as finding of metals imported from Tien Shan Mountains in Muradym 8) could therefore reflect a connection to the complex networks of the nomadic transmigration patterns characteristic of seasonal steppe population movements [see (2), figure 6.1, p. 205]. These movements, although dictated by the needs of the nomads and their animals, shaped the economic and social networks linking the outskirts of the steppe and facilitated the flow of goods between settled, semi-nomadic, and nomadic peoples. In contrast, all Cimmerians carried the Siberian genetic component. Both the PCA and f4 statistics supported their closer affinities to the Bronze Age western Siberian populations (including Karasuk) than to Srubnaya. It is noteworthy that the oldest of the Cimmerians studied here (cim357) carried almost equal proportions of Asian and West Eurasian components, resembling the Pazyryks, Aldy-Bel, and Iron Age individuals from Russia and Kazakhstan (12). The second oldest Cimmerian (cim358) was also the only one with both uniparental markers pointing toward East Asia. The Q1* Y chromosome sublineage of Q-M242 is widespread among Asians and Native Americans and is thought to have originated in the Altai Mountains (24). It has previously been identified in numerous ancient samples from Siberia, the Americas, and in representatives of the Siberian Bronze Age and nomadic populations (4, 24). This is the first indication that Cimmerians did not originate in the PCS region but were nomads tracing their origin to the Far East. The origins of Scythians of the western Pontic-Caspian steppe are difficult to resolve. We identified four different clusters within our geographically continuous sample set, which likely represent a varying gradient of different genetic components: the Northern cluster, SC, CC, and SE cluster. The latter was characterized by the presence of the NEN component representing local semi-nomadic Scythians with clear genetic uptake from the locals and possibly from other settlers such as the Greeks around the Black Sea region. Finally, the Sarmatians fell between all other nomads that form the bulk of the SC, suggesting that southern Urals is where the continuity of western nomads was sustained. Intragroup genetic diversity of the steppe populations The diversity of Srubnaya-Alakulskaya individuals was on par with that of Sarmatians and Scythians (table S10) and was also the highest among all published Bronze Age individuals. The Muradym 8/Kazburun 1 site is a unique cultural mixing zone with an unusual number of culturally distinct burials of two different traditions (Srubnaya-Alakulskaya), but the individuals are genetically uniform across the time span of the sites, suggesting that diffusion may have been the main mode of cultural dispersal in the LBA. Cimmerians resemble individuals within the SC and had higher diversity compared to that of the Srubnaya-Alakulskaya individuals. Despite clustering tightly on the PCA plot, the Cimmerians and Sarmatians had the highest pairwise mismatch estimates among the nomads studied (Fig. 1C and table S10). Although the Scythians showed high variation based on the PCA, their intragroup genetic diversity was comparatively lower than that of the Cimmerians and Sarmatians. This observation is best explained by a smaller ancestral Scythian population base in western Pontic-Caspian steppe than in Sarmatians of the southern Urals. When compared with published Early Sarmatians from the same region (12), the Sarmatians studied here seem to have remained genetically uniform through an approximately 300- to 500-year time span. This might suggest the genetic continuity of Sarmatians in the southern Urals despite a distinct cultural shift and a suspected earlier population replacement between Early and Middle/Late Sarmatians in the region. Thus, the observed high genetic diversity in the Sarmatians could result from a large effective population size rather than gene flow into the region.  Mutual relations and shared ancestry between steppe populations Our results suggest that the Cimmerians were largely similar to the more eastern Sarmatians (cim357) albeit with an increased amount of a Siberian (NEA) component (cim358 and cim359), thus representing a genetic link between the Iron Age people of the Kazakh steppe region. Similar to the cis-Uralic Srubnaya-Alakulskaya, Cimmerians were not direct ancestors of the Scythians in the Pontic-Caspian steppe; however, all those populations shared a common ancestral gene pool. Scythians who were thought to have displaced the Cimmerians from the Pontic-Caspian steppe region differed from both Cimmerians and the later Sarmatians. However, it is unclear whether the observed pattern resulted from replacement, since the genetic composition of Cimmerians may have undergone a temporal change, as witnessed by the observed temporal reduction of the NEA and the SEA components in more recent individuals. It has previously been noted that Scythians were not uniform across Eurasia, exhibiting distinct differences between Eastern/Western Eurasian groups (12). In contrast to the eastern steppe Scythians (Pazyryks and Aldy-Bel) that were closely related to Yamnaya, the western North Pontic Scythians were instead more closely related to individuals from Afanasievo and Andronovo groups. Some of the Scythians of the western Pontic-Caspian steppe lacked the SA and the East Eurasian components altogether and instead were more similar to a Montenegro Iron Age individual (3), possibly indicating assimilation of the earlier local groups by the Scythians. Finally, one individual from Nesterivka (scy011), which was the most recent of the Scythians, and possibly representing a transition between different groups, carried a distinct resemblance to the later steppe Sarmatians, corroborating the idea of a more “stable”/”common” steppe signature shared with all Sarmatians or reflecting physical/social connection to the southern Ural steppe zone maintained within the western nomadic world. Toward the end of the Scythian period (fourth century CE), a possible direct influx from the southern Ural steppe zone took place, as indicated by scy332. However, it is possible that this individual might have originated in a different nomadic group despite being found in a Scythian cultural context. This individual instead resembles, genetically, the early eastern Altai Scythians, albeit with even higher contribution of the NEA and the SEA genetic components. This result suggests the presence of a continuous connection between the Western fringes of the Scythian empire and central parts of Eurasia or maybe even the source region in the Altai Mountains. Such a connection could reflect communication associated with population exchange and internal high mobility among the Scythians. In conclusion, our genomic analyses revealed that both the Bronze Age and Iron Age were highly dynamic periods in the Pontic-Caspian steppe. The time span between 1800 BCE and 400 CE was characterized by mobility, population movements, and replacements, which shaped the complex demography of the region through time. Our results showed that the Western Eurasian steppe nomads were not direct descendants of the Bronze Age Srubnaya-Alakulskaya individuals but shared elements of common ancestry with contribution from different peoples. The early nomads could thus be referred to as a “cultural and chronological horizon” represented by various cultures of the Scythian-Siberian world that was not composed of a genetically homogeneous and/or isolated group. Quite the contrary is observed. We observe little evidence of mobility from the Far East, suggesting that the main source of most Western nomads is likely found in eastern Pontic-Caspian steppe and southern Urals. Thus, we propose that the region, similar to the so-called Mongolian steppe generator of peoples during the Middle Ages, served as the generator of the west nomadic peoples that sustained the western nomadic horizon in the Iron Age. Science Advances 03 Oct 2018: Vol. 4, no. 10, eaat4457 |
|
|
|
Post by Admin on Mar 31, 2019 17:51:25 GMT
 A TRIBE who swept into Europe thousands of years ago and whose descendants wiped out ancient Britons could be the most violent and aggressive society ever, it was claimed. The Yamnaya were a group of livestock herders who lived north of the Black Sea and in the Caucasus mountains in modern day Russia and Ukraine. The group and their descendants arrived in Europe between 4000 and 5000 years ago, New Scientist reports.bThe migrants’ ultimate source was a group of livestock herders called the Yamnaya who occupied the Eurasian steppe north of the Black Sea and the Caucasus mountains. Britain wasn’t their only destination. Between 5000 and 4000 years ago, the Yamnaya and their descendants colonised swathes of Europe, leaving a genetic legacy that persists to this day. Their arrival coincided with profound social and cultural changes. Burial practices shifted dramatically, a warrior class appeared, and there seems to have been a sharp upsurge in lethal violence. “I’ve become increasingly convinced there must have been a kind of genocide,” says Kristian Kristiansen at the University of Gothenburg, Sweden. As he and others piece together the story, one question resounds: were the Yamnaya the most murderous people in history? 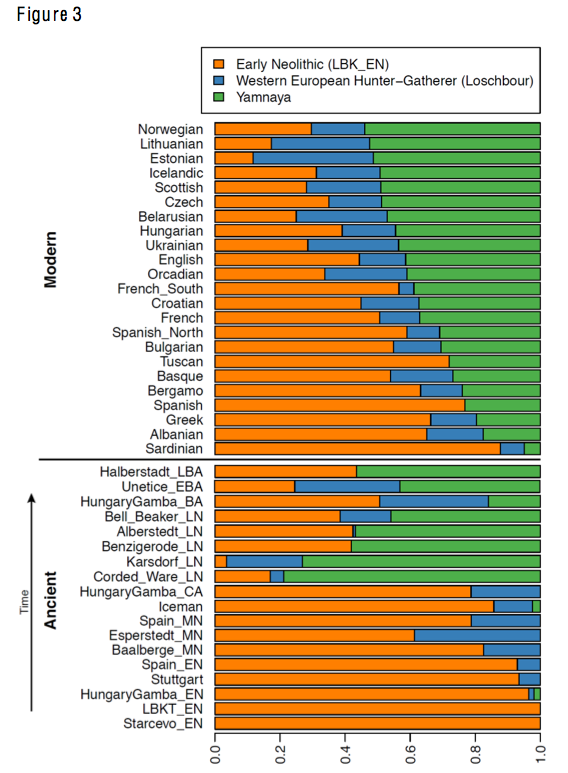 At the time Europe had been ravaged by disease, eroding the population’s ability to resist the powerfully built and aggressive Yamnaya horseback warriors. They overwhelmed smaller Europeans and the Yamnaya culture as well as their DNA soon spread throughout the continent. According archaeologist Kristian Kristiansen, after they brought with their brutal practices there appears to have been a sharp upsurge in lethal violence. Such was the extent of their brutality that he began to consider whether they were the most murderous people in history. “I’ve become increasingly convinced there must have been a kind of genocide,” said Professor Kristiansen. 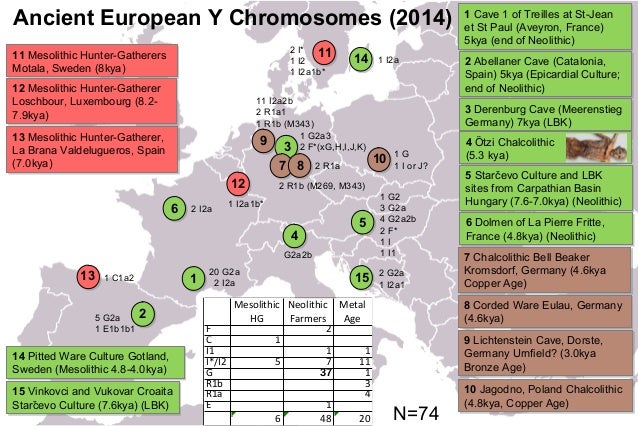 He said the Yamnaya lived mostly on meat and milk products which made them “healthier and probably physically quite strong”. It has been claimed that when the Yamnaya invaded they massacred all the males and impregnated the women in order to rapidly further their bloodlines. This would have caused panic and resistance in Neolithic societies struggling to hold back the Yamnaya. Graves show the remains of the Eulau who were captured and massacred in retaliation. It has been claimed that the Yamnaya are the missing third genetic component in Europeans.  A team led by Professor David Reich and his colleagues extracted DNA from remains found at archaeological sites around the continent, the BBC reported. Their research showed that 7,000-8,000 years ago, a closely related group of early farmers moved into Europe from the Near East. This group interbred with indigenous hunter-gatherers that they encountered as they spread around the continent and eventually the two groups melded. |
|
|
|
Post by Admin on Nov 13, 2019 17:53:17 GMT
Abstract Hun, Avar and conquering Hungarian nomadic groups arrived to the Carpathian Basin from the Eurasian Steppes and significantly influenced its political and ethnical landscape, however their origin remains largely unknown. In order to shed light on the genetic affinity of above groups we have determined Y chromosomal haplogroups and autosomal loci, suitable to predict biogeographic ancestry, from 49 individuals, supposed to represent the power/military elit. Haplogroups from the Hun-age are consistent with Xiongnu ancestry of European Huns. Most of the Avar-age individuals carry east Eurasian Y haplogroups typical for modern north-eastern Siberian and Buryat populations and their autosomal loci indicate mostly un-admixed Asian characteristics. In contrast the conquering Hungarians seem to be a recently assembled population incorporating un-admixed European, Asian as well as admixed components. Their heterogeneous paternal and maternal lineages indicate similar supposed phylogeographic origin of males and females, derived from Central-Inner Asian and European Pontic Steppe sources.  Population history of the Carpathian Basin was profoundly determined by the invasion of various nomadic groups from the Eurasian Steppes during the Middle Ages. Between 400–453 AD the Huns held possession of the region and brought about a major population reshuffling all over Europe. Recent genetic data connect European Huns to Inner Asian Xiongnus1, but genetic data from Huns of the Carpathian Basin have not been available yet, since Huns left just sporadic lonely graves in the region, as they stayed for short period. We report three Y haplogroups (Hg) from Hun age remains, which possibly belonged to Huns based on their archaeological and anthropological evaluation. From 568 AD the Avars established an empire in the region lasting nearly for 250 years, until they were defeated by the Franks and Bulgars in 803, then their steppe-empire ended around 822 AD. In its early stage the Avar Khaganate controlled a large territory expanding from the Carpathian Basin to the Pontic-Caspian Steppes and dominated numerous folks including Onogur-Bulgars, which fought their independence in the middle 7th century and established the independent Magna Bulgaria state. Then the Avar Khaganate was shrunken, its range well corresponding to that of the succeeding Hungarian Kingdom. The Avars arrived in multiple waves into the Carpathian Basin and the Avar period left a vast archeological legacy with more than 80 thousand excavated graves in present-day Hungary. The Avar age remains are anthropologically extremely heterogeneous, with considerable proportion of Mongoloid and Europo-Monoloid elements reaching 20–30% on the Great Hungarian Plain2, attesting that the Carpathian Basin witnessed the largest invasion of people from Asia during this period. Most individuals buried with rich grave goods show Mongoloid characters indicating inner Asian origin of the Avar elite, which is also supported by their artifact types, titles (e.g. khagan) and institutions recognized to be derived from Inner Asian Rourans. From the Avar period only a few mitochondrial DNA (mtDNA) data are available from two micro-regions3,4, which showed 15.3% and 6.52% frequency of East Eurasian elements. A recent manuscript described 23 mitogenomes from the 7th-8th century Avar elite group5 and found that 64% of the lineages belong to East Asian haplogroups (C, D, F, M, R, Y and Z) with affinities to ancient and modern Inner Asian populations corroborating their Rouran origin. Though the Avar Khaganate ceased to exist around 822 AD, but its population survived and were incorporated into the succeeding Hungarian state6. It is relevant to note that none of the Hungarian medieval sources know about Avars7, probably because they were not distinguished from the Huns as many foreign medieval sources also identified Avars with Huns, for example the Avars who were Christianized and became tax-payer vassals of the Eastern Frankish Empire were called as Huns in 8718.  Presence of the Hungarians in the Carpathian Basin was documented from 862 AD and between 895–905 they took full command of the region. The Hungarians formed a tribal union but arrived in the frame of a strong centralized steppe-empire under the leadership of prince Álmos and his son Árpád, who were known to be direct descendants of the great Hun leader Attila, and became founders of the Hungarian ruling dynasty and the Hungarian state. The Hungarian Great Principality existed in Central Europe from ca. 862 until 1000, then it was re-organized as a Christian Kingdom by King István I the Saint who was the 5th descendant of Álmos9. Our recent analysis of conquering Hungarian (hence shortened as Conqueror) mitogenomes revealed that the origin of their maternal lineages can be traced back to distant parts of the Eurasian steppe10. One third of the maternal lineages were derived from Central-Inner Asia and their most probable ultimate sources were the Asian Scythians and Asian Huns, while the majority of the lineages most likely originated from the Bronze Age Potapovka-Poltavka-Srubnaya cultures of the Pontic-Caspian steppe. Population genetic analysis indicated that Conquerors had closest connection to the Onogur-Bulgar ancestors of Volga Tatars. 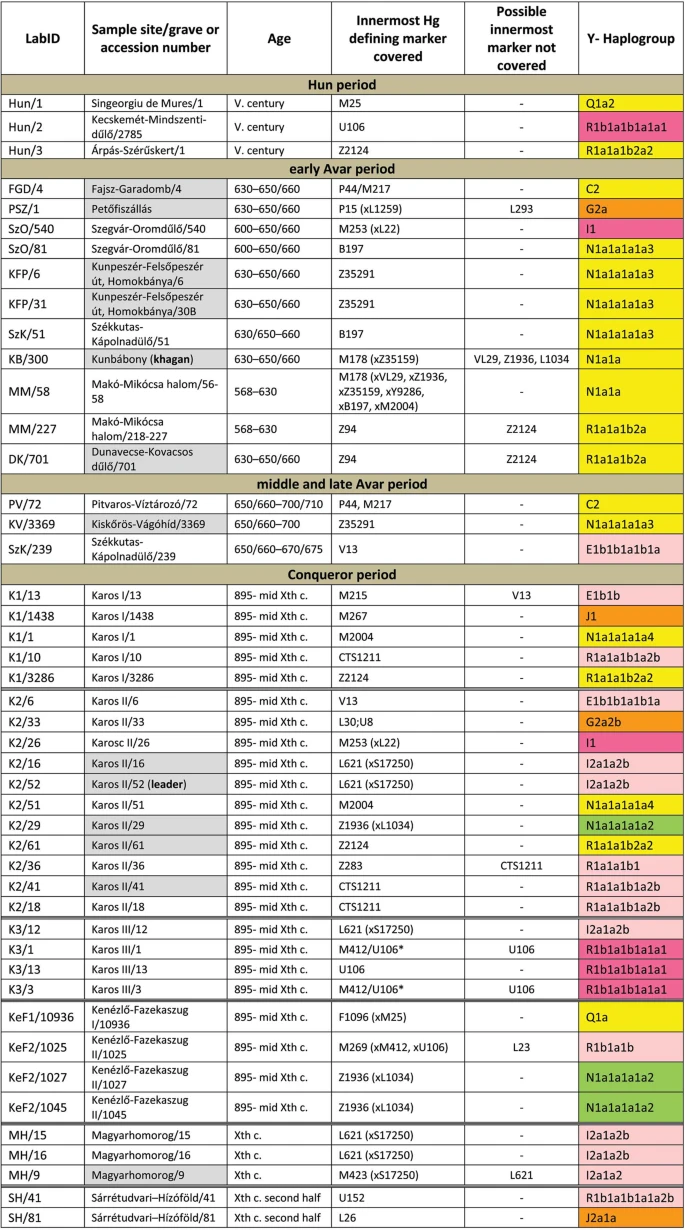 Figure 1 Although mtDNA data can be informative, since as far as we know none of the above nomadic invasions were pure military expeditions in which raiding males took local women, but entire societies with both men and women migrated together3, however nomadic societies were patrilinealy organized, thus Y-chromosome data are expected to provide more relevant information about their structure and origin than mtDNA. So far 6 Y-chromosome Hg-s have been published from the Conquerors11; which revealed the presence of N1a1- M46 (previously called Tat or N1c), in two out of 4 men, while12 detected two R1b-U106 and two I2a-M170 Hg-s. In order to generate sufficient data for statistical evaluation and compare paternal and maternal lineages of the same population, we have determined Y-Hg-s from the same cemeteries whose mtDNA Hg-s were described in10. As most Hungarian medieval chronicles describe the conquer as the “second incoming of the Hungarians”7, genetic relations may exist between consecutive nomadic groups, first of all between elite groups. As the studied Conqueror cemeteries represent mainly the Conqueror elite, we supplemented our Y-Hg studies with samples from the preceding Avar power elite/millitary elite groups and available samples from the Hun period to test possible genetic relations. Avar military leader graves are distinguished by high-value prestige artifacts: belt decorated with gold or silver mounts, decorated horse harness, saddle, sword, bow and quiver with arrowheads, gold decorated plates, silver or gold earrings (Supplementary Information). Conqueror leader graves contain characteristic partial horse burial with horse cranium and leg bones or symbolic horse burial with adorned harness, saddle and stirrup. Elite grave goods often include precious metal jewels, belt with impressed metal belt-buckles, metal-plated sabretache and weaponry; arrowheads, quiver and bow10,12. ry. |
|
|
|
Post by Admin on Nov 14, 2019 3:56:53 GMT
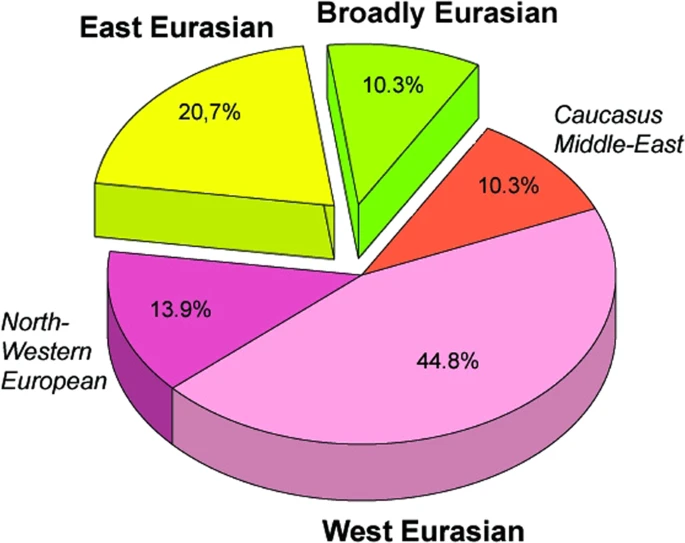 Figure 2 East Eurasian Hg-s Hg Q1a2- M25 is very rare in Europe, where it has highest frequency among Seklers (a Hungarian speaking ethnic group in Transylvania) according to Family Tree DNA database. Ancient samples with Hg Q1a2- M25 are known from the Bronze Age Okunevo and Karasuk cultures, as well as Middle Age Tian Shan Huns and Hunnic-Sarmatians17 implying possible Hunnic origin of this lineage in Europe, which is confirmed by the Hg of our Hun/1 sample, derived from Transylvania. One Conqueror sample KEF1/10936 belongs to Q1a- F1096, possibly to the Q1a1-F746 sister clade of Q1a2- M25, which was not tested in our experiment. Q1a1 is present in East Asia; Mongolia, Japan, China, Korea and its presence in the Kenézlő (KEF) graveyard imply a common substrate of Huns and Conquerors. Hun/3 belongs to Hg R1a1a1b2a2- Z2124, a subclade of R1a1a1b2-Z93, the east Eurasian subbranch of R1a. Today Z2124 is most frequent in Kyrgyzstan and Afghanistan, but is also widespread among Karachai-Balkars and Baskhirs18. Z2124 was widespread on the Bronze Age steppe, especially in the Afanasievo and Sintashta cultures19 and R1a detected in Xiongnus20,21 very likely belong to the same branch. Two samples from the Karos Conqueror cemeteries (K1/3286 and K2/61) were also classified as R1a-Z2124 and two Avar age individuals (DK/701 and MM/227) belong to the same R1a1a1b2a-Z94 branch but marker Z2124 was not covered in latter samples. Two Avar samples belonged to Hg C2-M217, which is found mostly in Central Asia and Eastern Siberia. Presence of this Hg had also been detected in the Conquerors, as the Karos2/60 individual belongs to C2 (Fóthi unpublished). All N-Hg-s identified in the Avars and Conquerors belonged to N1a1a-M178. We have tested 7 subclades of M178; N1a1a2-B187, N1a1a1a2-B211, N1a1a1a1a3-B197, N1a1a1a1a4-M2118, N1a1a1a1a1a-VL29, N1a1a1a1a2-Z1936 and the N1a1a1a1a2a1c1-L1034 subbranch of Z1936. The European subclades VL29 and Z1936 could be excluded in most cases, while the rest of the suclades are prevalent in Siberia22 from where this Hg dispersed in a counter-clockwise migratory route to Europe23. Avar sample MM/58, did not go into any of the tested M178 subclades, while only N1a1a2 could be excluded for the KB/300 Avar khagan due to low coverage. All the 5 other Avar samples belonged to N1a1a1a1a3-B197, which is most prevalent in Chukchi, Buryats, Eskimos, Koryaks and appears among Tuvans and Mongols with lower frequency22. By contrast two Conquerors belonged to N1a1a1a1a4-M2118, the Y lineage of nearly all Yakut males, being also frequent in Evenks, Evens and occurring with lower frequency among Khantys, Mansis and Kazakhs. Broadly Eurasian Hg-s Three Conqueror samples belonged to Hg N1a1a1a1a2-Z1936, the Finno-Permic N1a branch, being most frequent among northeastern European Saami, Finns, Karelians, as well as Komis, Volga Tatars and Bashkirs of the Volga-Ural region. Nevertheless this Hg is also present with lower frequency among Karanogays, Siberian Nenets, Khantys, Mansis, Dolgans, Nganasans, and Siberian Tatars22. 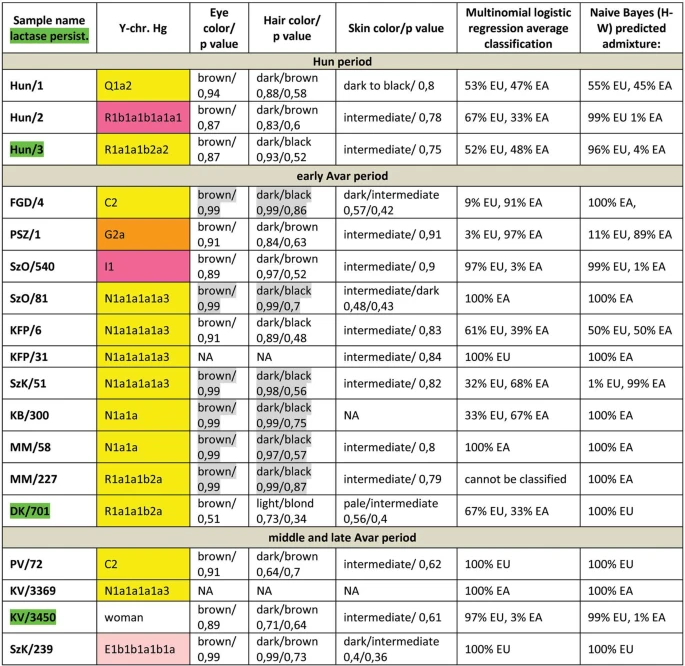 Figure 3 West Eurasian Hg-s The west Eurasian R1a1a1b1a2b-CTS1211 subclade of R1a is most frequent in Eastern Europe especially among Slavic people. This Hg was detected just in the Conqueror group (K2/18, K2/41 and K1/10). Though CTS1211 was not covered in K2/36 but it may also belong to this sub-branch of Z283. Hg I2a1a2b-L621 was present in 5 Conqueror samples, and a 6th sample form Magyarhomorog (MH/9) most likely also belongs here, as MH/9 is a likely kin of MH/16 (see below). This Hg of European origin is most prominent in the Balkans and Eastern Europe, especially among Slavic speaking groups. It might have been a major lineage of the Cucuteni-Trypillian culture and it was present in the Baden culture of the Calcholitic Carpathian Basin24. I1- M253, identified from one Conqueror sample is a northern European Hg found mostly in Scandinavia and Finland and might have originated from this region during the Mesolithic. It has somewhat similar distribution to R1b-U106 associated with Germanic speaking populations. Three out of 4 samples in the small Karos3 cemetery belonged to Hg R1b1a1b1a1a1-U106 setting apart this cemetery from all other groups, except for the Hun/2 sample which is the only other one with this Hg. Hg U106 is considered a “Germanic” branch as it is most significant today in Germany, Scandinavia, and Britain, and rare in Eastern Europe (Supplementary Table S4). Its ancestral branch Hg R1b1a1b-M262 is assumed to have emerged in the Pontic-Caspian Steppe and arrived to Europe with Bronze Age migrations25. Its presence in Hun and Conqueror samples may derive from Goths, Gepids or other German allies of the Huns. We detected R1b1a1b1a1a2b-U152 in one sample from the Sárrétudvari Conqueror cemetery, which represent rather commoners than the elite. U152 is the Italo-Celtic R1b branch, concentrated around the Alps, and which was present in the Carpathian Basin before the conquer, so did not necessarily arrived with the Conquerors. The mediterranean haplogroup E1b1b1a1b1a-V13 was detected in an Avar (SzK/239) and a Conqueror (K2/6) sample, while this marker was not covered in another sample (K1/13, E1b1b- M215). This Hg originated in the Middle East and migrated to the Balkans and Western Asia during the Bronze Age. The PSZ/1 Avar leader belongs to Hg G2a-P15, while K2/33 to the G2a2b-L30 subbranch. Hg G2a is originated from Anatolia/Iran, now it is most common in the Caucasus region and its arrival to Europe is associated with the spread of Neolithic farmers26. One Conqueror sample belongs to Hg J1-M267 and another to J2a1a-L26. Both J1 and J2 lineages are most frequent around the Middle East-Caucasus and probably originated from this region27, then expanded with pastorists prior to the Neolithic. |
|

