|
|
Post by Admin on Feb 16, 2020 23:33:09 GMT
 Fig. 3 Genetic relationship between Srubnaya-Alakulskaya population and Iron Age nomads. Visual summary of f4 statistics of a form f4(YRI, TestPop, Pop X, Srubnaya), where TestPop is (A) Cimmerian; (B) Scythian and (C) Sarmatian. (D) Admixture selection: Our ancient individuals and Iron Age individuals from closely related populations (K = 15). K = 15 was chosen to display since it shows SA (lilac) and NEA (dark green) components in addition to the other components. Srubnaya-Alakulskaya population is named as Srubnaya in the panels. IA, Iron Age (fig. S10). In the seventh century BCE, the Scythians appeared in the North Pontic region. Iron Age western Scythians displayed slightly higher intragroup diversity compared to that of the Bronze Age groups and formed four discrete clusters including NE, SE, SC, and CC clusters (Fig. 1C and table S10). Scythians belonging to the SE cluster were closer to Hungarian Bronze Age and Iron Age individuals including Vatya and Maros. The NE Scythian cluster fell close to the Iron Age individuals from modern-day Montenegro and Sweden (fig. S9). The SC Scythians further grouped with Early Sarmatians (12) and the Iron Age Scythian from modern-day Hungary (23). It has been hypothesized by Terenozhkin that Scythians reached the Pontic-Caspian steppe region from Central Asia (for example, from Andronovo) (7). To formally test this hypothesis, we calculated f4 statistics in the form of f4(Yoruba, Scythian, Srubnaya, BronzeAgeX), where “BronzeAgeX” refers to either Andronovo, Sintashta, or Afanasievo. This analysis revealed that the western Scythians (tested either as a single population or four different clusters) formed a clade together with Andronovo and two other Andronovo-originated populations to the exclusion of Srubnaya-Alakulskaya from the southern Urals (Fig. 3B and table S14), supporting the Central Asian origin of the western Scythians. Furthermore, the western Scythians shared more drift with Andronovo, Afanasievo, Sintashta, and Mezhovskaya to the exclusion of Yamnaya (table S15). While the eastern Scythians from an earlier study (12) formed a clade together with Srubnaya-Alakulskaya to the exclusion of Yamnaya, the western Scythians of the present study did not show this pattern. Only when compared with Karasuk and Okunevo, Yamnaya seemed closer (although scy332 is an exception), which is in line with a shared steppe origin of the Yamanya and the western nomads (table S15). ADMIXTURE analysis at K = 15 revealed that the Scythians belonging to the NE cluster had a more visible West Eurasian component that is highest in the northern populations, while Scythians that belong to the SE cluster had a more visible NEN genetic component, most commonly found in Neolithic Anatolian individuals. The NEN component was also found in individuals falling into the CC, who additionally carried an SA component (“lilac”) shared with the Cimmerians, Sarmatians, and two more Scythian individuals (scy193 and scy006). Scy332 had a SEA (“light blue”) component and was most similar to the Karasuk individuals (Fig. 3 and fig. S10, A and B). The five Sarmatians grouped close together in the SC (Fig. 1C), but they also had the highest pairwise mismatch estimates compared to other Iron Age nomads, suggesting a larger effective population size (table S10). F4(Yoruba, Sarmatians, PopulationX, Cimmerian), where the PopulationX is Yamnaya, Srubnaya, or Karasuk, revealed that Sarmatians formed a clade together with all these three populations to the exclusion of Cimmerians (table S18). Multiway f4 statistics testing the relationships between Srubnaya/-Alakulskaya, Cimmerians, and Scythians revealed that both Scythians and Cimmerians formed a clade together with Srubnaya/-Alakulskaya to the exclusion of the other population and that Srubnaya/-Alakulskaya was closer to Scythians among the two (Fig. 3C and table S17). The results point to the presence of a deep shared ancestry of all Iron Age nomadic groups associated with Bronze Age populations of the steppe, which, however, is not equivalent with a direct genetic continuity between Srubnaya-Alakulskaya and the western Scythians. DISCUSSION The origins of the four steppe populations The Bronze Age Srubnaya-Alakulskaya individuals from Kazburun 1/Muradym 8 presented genetic similarities to the previously published Srubnaya individuals. However, in f4 statistics, they shared more drift with representatives of the Andronovo and Afanasievo populations compared to the published Srubnaya individuals. Those apparently West Eurasian people lacked significant Siberian components (NEA and SEA) in ADMIXTURE analyses but carried traces of the SA component that could represent an earlier connection to ancient Bactria. The presence of an SA component (as well as finding of metals imported from Tien Shan Mountains in Muradym 8) could therefore reflect a connection to the complex networks of the nomadic transmigration patterns characteristic of seasonal steppe population movements [see (2), figure 6.1, p. 205]. These movements, although dictated by the needs of the nomads and their animals, shaped the economic and social networks linking the outskirts of the steppe and facilitated the flow of goods between settled, semi-nomadic, and nomadic peoples. In contrast, all Cimmerians carried the Siberian genetic component. Both the PCA and f4 statistics supported their closer affinities to the Bronze Age western Siberian populations (including Karasuk) than to Srubnaya. It is noteworthy that the oldest of the Cimmerians studied here (cim357) carried almost equal proportions of Asian and West Eurasian components, resembling the Pazyryks, Aldy-Bel, and Iron Age individuals from Russia and Kazakhstan (12). The second oldest Cimmerian (cim358) was also the only one with both uniparental markers pointing toward East Asia. The Q1* Y chromosome sublineage of Q-M242 is widespread among Asians and Native Americans and is thought to have originated in the Altai Mountains (24). It has previously been identified in numerous ancient samples from Siberia, the Americas, and in representatives of the Siberian Bronze Age and nomadic populations (4, 24). This is the first indication that Cimmerians did not originate in the PCS region but were nomads tracing their origin to the Far East. The origins of Scythians of the western Pontic-Caspian steppe are difficult to resolve. We identified four different clusters within our geographically continuous sample set, which likely represent a varying gradient of different genetic components: the Northern cluster, SC, CC, and SE cluster. The latter was characterized by the presence of the NEN component representing local semi-nomadic Scythians with clear genetic uptake from the locals and possibly from other settlers such as the Greeks around the Black Sea region. Finally, the Sarmatians fell between all other nomads that form the bulk of the SC, suggesting that southern Urals is where the continuity of western nomads was sustained. Intragroup genetic diversity of the steppe populations The diversity of Srubnaya-Alakulskaya individuals was on par with that of Sarmatians and Scythians (table S10) and was also the highest among all published Bronze Age individuals. The Muradym 8/Kazburun 1 site is a unique cultural mixing zone with an unusual number of culturally distinct burials of two different traditions (Srubnaya-Alakulskaya), but the individuals are genetically uniform across the time span of the sites, suggesting that diffusion may have been the main mode of cultural dispersal in the LBA. Cimmerians resemble individuals within the SC and had higher diversity compared to that of the Srubnaya-Alakulskaya individuals. Despite clustering tightly on the PCA plot, the Cimmerians and Sarmatians had the highest pairwise mismatch estimates among the nomads studied (Fig. 1C and table S10). Although the Scythians showed high variation based on the PCA, their intragroup genetic diversity was comparatively lower than that of the Cimmerians and Sarmatians. This observation is best explained by a smaller ancestral Scythian population base in western Pontic-Caspian steppe than in Sarmatians of the southern Urals. When compared with published Early Sarmatians from the same region (12), the Sarmatians studied here seem to have remained genetically uniform through an approximately 300- to 500-year time span. This might suggest the genetic continuity of Sarmatians in the southern Urals despite a distinct cultural shift and a suspected earlier population replacement between Early and Middle/Late Sarmatians in the region. Thus, the observed high genetic diversity in the Sarmatians could result from a large effective population size rather than gene flow into the region. Mutual relations and shared ancestry between steppe populations Our results suggest that the Cimmerians were largely similar to the more eastern Sarmatians (cim357) albeit with an increased amount of a Siberian (NEA) component (cim358 and cim359), thus representing a genetic link between the Iron Age people of the Kazakh steppe region. Similar to the cis-Uralic Srubnaya-Alakulskaya, Cimmerians were not direct ancestors of the Scythians in the Pontic-Caspian steppe; however, all those populations shared a common ancestral gene pool. Scythians who were thought to have displaced the Cimmerians from the Pontic-Caspian steppe region differed from both Cimmerians and the later Sarmatians. However, it is unclear whether the observed pattern resulted from replacement, since the genetic composition of Cimmerians may have undergone a temporal change, as witnessed by the observed temporal reduction of the NEA and the SEA components in more recent individuals. It has previously been noted that Scythians were not uniform across Eurasia, exhibiting distinct differences between Eastern/Western Eurasian groups (12). In contrast to the eastern steppe Scythians (Pazyryks and Aldy-Bel) that were closely related to Yamnaya, the western North Pontic Scythians were instead more closely related to individuals from Afanasievo and Andronovo groups. Some of the Scythians of the western Pontic-Caspian steppe lacked the SA and the East Eurasian components altogether and instead were more similar to a Montenegro Iron Age individual (3), possibly indicating assimilation of the earlier local groups by the Scythians. Finally, one individual from Nesterivka (scy011), which was the most recent of the Scythians, and possibly representing a transition between different groups, carried a distinct resemblance to the later steppe Sarmatians, corroborating the idea of a more “stable”/”common” steppe signature shared with all Sarmatians or reflecting physical/social connection to the southern Ural steppe zone maintained within the western nomadic world. Toward the end of the Scythian period (fourth century CE), a possible direct influx from the southern Ural steppe zone took place, as indicated by scy332. However, it is possible that this individual might have originated in a different nomadic group despite being found in a Scythian cultural context. This individual instead resembles, genetically, the early eastern Altai Scythians, albeit with even higher contribution of the NEA and the SEA genetic components. This result suggests the presence of a continuous connection between the Western fringes of the Scythian empire and central parts of Eurasia or maybe even the source region in the Altai Mountains. Such a connection could reflect communication associated with population exchange and internal high mobility among the Scythians. In conclusion, our genomic analyses revealed that both the Bronze Age and Iron Age were highly dynamic periods in the Pontic-Caspian steppe. The time span between 1800 BCE and 400 CE was characterized by mobility, population movements, and replacements, which shaped the complex demography of the region through time. Our results showed that the Western Eurasian steppe nomads were not direct descendants of the Bronze Age Srubnaya-Alakulskaya individuals but shared elements of common ancestry with contribution from different peoples. The early nomads could thus be referred to as a “cultural and chronological horizon” represented by various cultures of the Scythian-Siberian world that was not composed of a genetically homogeneous and/or isolated group. Quite the contrary is observed. We observe little evidence of mobility from the Far East, suggesting that the main source of most Western nomads is likely found in eastern Pontic-Caspian steppe and southern Urals. Thus, we propose that the region, similar to the so-called Mongolian steppe generator of peoples during the Middle Ages, served as the generator of the west nomadic peoples that sustained the western nomadic horizon in the Iron Age. Science Advances 03 Oct 2018: Vol. 4, no. 10, eaat4457 |
|
|
|
Post by Admin on Feb 17, 2020 21:01:55 GMT
Abstract One of the best documented Indo-European civilizations that inhabited Bulgaria is the Thracians, who lasted for more than five millennia and whose origin and relationships with other past and present-day populations are debated among researchers. Here we report 25 new complete mitochondrial genomes of ancient individuals coming from three necropolises located in different regions of Bulgaria – Shekerdja mogila, Gabrova mogila and Bereketska mogila – dated to II-III millennium BC. The identified mtDNA haplogroup composition reflects the mitochondrial variability of Western Eurasia. In particular, within the ancient Eurasian genetic landscape, Thracians locate in an intermediate position between Early Neolithic farmers and Late Neolithic-Bronze Age steppe pastoralists, supporting the scenario that the Balkan region has been a link between Eastern Europe and the Mediterranean since the prehistoric time. Spatial Principal Component Analysis (sPCA) performed on Thracian and modern mtDNA sequences, confirms the pattern highlighted on ancient populations, overall indicating that the maternal gene pool of Thracians reflects their central geographical position at the gateway of Europe. Introduction Bulgaria is situated in the eastern part of the Balkan Peninsula, at the connection point between Southeastern Europe, Eurasian steppe, Anatolia and the Aegean islands. The presence of modern humans in this region is attested starting from 40 kya by the Paleolithic series at Bacho Kiro and Temnata Dupka Caves1,2,3. Some archaeological sites associated with early farmers, as well as the earliest evidence of copper metallurgy in Europe, indicates that this area played a significant role both in the Neolithic and in the Metal Ages4. One of the best documented Indo-European civilizations that inhabited Bulgaria consists in the Thracians, whose cultural legacy is still evident in the modern society. Different theories have been historically proposed about the origin of the Thracians. Today it is assumed that the Thracian culture emerged and formed in the early Bronze Age5,6,7, a period characterized by strong cultural changes and movements of people westward from the Steppe8. During the 5th and 4th millennium BCE, the inhabitants of the eastern region of Balkans were organized in different groups of indigenous people that, over time, were named under the single ethnonym of “Thracians”9,10,11. According to historical and archaeological sources, the Thracian culture flourished during the 2nd and 3rd millennia BCE12,13. The rich cultural and historical heritage, represented by fortresses and necropolises, as well as by the world-famous Panagyurishte, Valchitran, Lukovit and Rogozen treasures, dates back to this period. In the later periods, several populations (Greeks, Macedonians, Slavs and proto-Bulgarians) arrived in the Balkans, reaching the lands occupied by Thracians and mixing with them, thus influencing their cultural and biological identity11. Genetic analyses on both autosomal variations14 and uniparental genetic markers15,16,17 of present-day Bulgarians, locate them between Eastern European and Mediterranean populations, with a particular affinity to the neighboring groups from Greece and the Balkans16. In addition, the Bulgarian maternal genetic pool particularly suggests a major Western Eurasian origin, tracing their ancestry to lineages that witness a complex genetic structure of the region today and reflect different peopling and admixture events from the Upper Paleolithic to the onset of the Neolithic and Post-Neolithic in Europe15. Recent genome-wide ancient DNA (aDNA) based studies on Southeastern Europe, have shown that Neolithic population from present-day Bulgaria was closely related with the northwestern-Anatolian-Neolithic ancestry that signals the spread of Early farmers across Europe, except for the individuals lived in the mid-sixth millennium BC in Malak Preslavets, who revealed a significantly higher level of hunter-gatherer-related ancestry than the other Balkan Neolithic individuals18. Starting from the early 3rd millennium BCE, migrations from the adjacent Pontic-Caspian and Eurasian steppe also played an important role in the transformation of the European genetic landscape, and the contribution of Steppe ancestry to Southeastern European populations increased particularly during the Bronze Age18. Although our understanding of the population and cultural dynamics occurred in the (pre-) history of Balkan Peninsula is starting to be increasingly elucidated, the genetic details on the local civilization remain unknown and this information is only partly available for the ancient (proto-) Bulgarian people18,19. We now have the opportunity to investigate the genetic structure of the Thracians, an ancient people that lasted for more than five millennia and whose origin and relationships with other past and present-day populations are still debated among researchers. 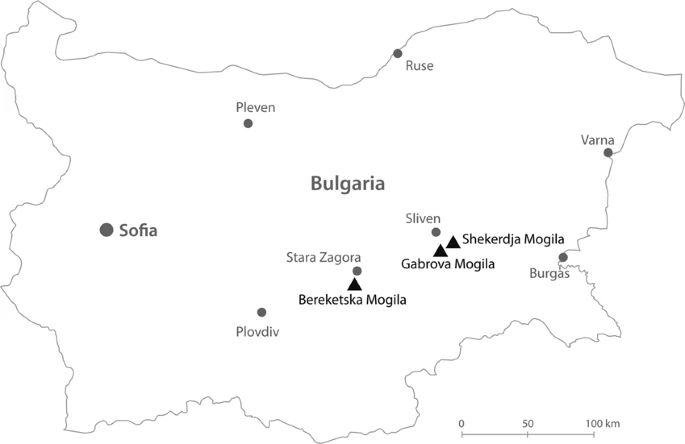 Figure 1 Geographical location of the necropolises in Bulgaria. Black triangles specifically indicate the locations of the considered archaeological sites while the grey dots refer to current Bulgarian provinces. The map is plotted using www.freepik.com/, processed with Adobe Illustrator CS6 and modified with Photoshop CS6 (2012) by Dimitar Spassov - web developer (dimitarspassov@gmail.com) and Desislava Nesheva. Image is attributed to valeria_aksakova/Freepik. To investigate the genetic structure and population history of this ancient civilization, we analyzed 25 complete mitochondrial genomes from three Thracian necropolises (Fig. 1 and Table 1) along with modern and ancient European data. The characterization of the Thracian mitochondrial DNA (mtDNA) variability may have important implications for understanding the dynamics of interaction between Eastern Europe and the Mediterranean, and will also contribute to better clarify the genetic evolution of European populations and the origin of contemporary Bulgarian gene pool. |
|
|
|
Post by Admin on Feb 18, 2020 21:00:12 GMT
Results We successfully reconstructed complete or almost entire mitochondrial genomes for 26 individuals, 3 from Shekerdja mogila, 1 from Gabrova mogila and 22 from Bereketska mogila (Table 2). All the resulted sequences reach the standard quality requested to guaranty the reliability of the NGS data; CtoT patterns range between 20% to 46%, average fragment size vary from 44.4 base pair (bp) to 67.4 bp and no significant levels of present-day human contamination were detected (Table 2). Only one sample, BM-51, showed a high level of contamination and was not considered in the following statistical analyses. The direct radiocarbon dating performed on the samples BM 44, SM 8.1 and GM 30.3 placed the remains at II-III millennium BC (CEDAD, Centro di DAtazione e Diagnostica, Univerità del Salento, Italy) (Table 1), that corresponds to the age estimated according to the archaeological record. The mtDNA sequences obtained were assigned to 21 different haplogroups, representative of the mitochondrial variability of Western Eurasia (Table 2 and Supplementary Table S1). Phylogenetic links between haplotypes of the Thracian samples and comparison ancient data are shown in the Median Joining Network (Fig. 2). Most of the Thracian individuals belong to sub-lineages of the macro-haplogroup H, which accounts for an overall frequency of 33%. This is the most frequent mitochondrial lineage in present-day Europe, representing over 40% of the total mtDNA variability20. Its frequency observed in the Thracians samples is almost similar to the frequency in contemporary European population. Two individuals belong to haplogroup HV, an ancient European lineage likely originating in the Mediterranean region during the Last Glacial Maximum (LGM)21. In ancient samples, HV has been identified in one Mesolithic specimen from Sicily22 and in early Neolithic remains from Spain23, Germany8 and Russia18,24; Mathieson et al.18 reported a HV haplotype in one sample from Serbia dating from 5800 BCE. Moreover, haplogroup HV was observed in Copper Age specimens from Scotland, Hungary and Germany25 and in Hungarian and Israeli samples from the Chalcolithic period26,27 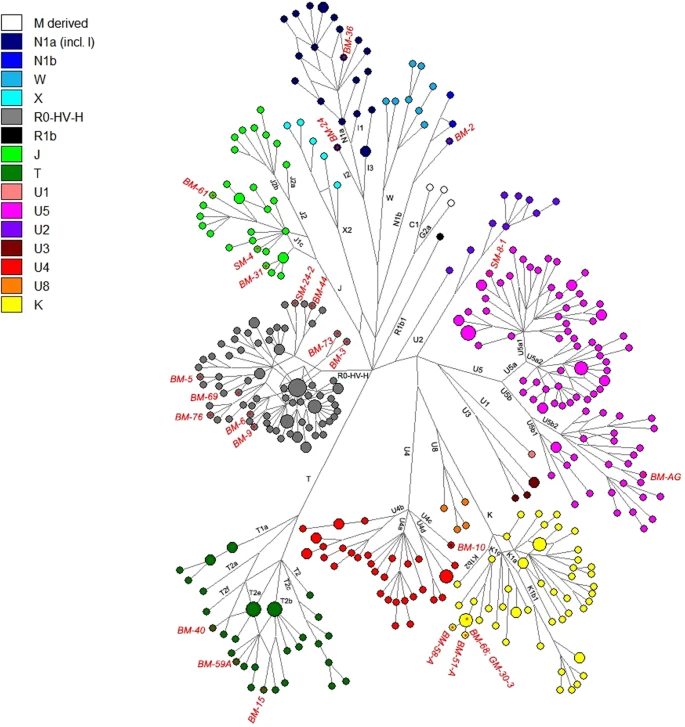 Figure 2 We found four individuals belonging to haplogroup K1c (GM-30.3, BM-51A, BM-58A and BM-68). All the haplotypes contain the expected K1c defining variants with the following private polymorphisms: GM-30.3, 309.1T, 310C, 7441T and 16519C; BM-51A, 16519C; BM-58A, 310C, 513.1CA and 16519C; BM-68, 5297T and 16519C. Nowadays the highest observed European frequency of the lineage K is in Bulgaria (13.3%)28 and K1c is particularly common in Slavic-speaking countries. In ancient populations, the haplogroup K1c has been identified in six hunter-gatherers dated before the arrival of farming (one in Romania, three in Serbia18 and two in Greece29), in two Bronze-Age individuals from Hungary and Bulgaria18,30,31 and in two Central-Europe farmers associated with the Bell-Beaker culture25,32,33. The phylogenetic network analysis (Fig. 2) reveals that the detected K1c haplotypes in Thracians are closely related to hunter-gatherers from Iron Gates and Bronze Age individuals from Bulgaria and Hungary. Three samples belong to haplogroup J1c (SM-4, BM-31 and BM-61). The SM-4 individual shows three personal transitions previously identified at positions 199C, 8730G and 13928A, and a private mutation at 13686G. The haplotypes of samples BM-31 and BM-61 fall within the sub-haplogropus J1c9 and J1c6, respectively. Currently, J1c, which dates to ∼16 ka ago, is found mainly in Europe, especially in Central Europe, Balkans and Ukraine, where it encompasses almost 80% of total J1 lineages. Pala et al.34 suggested that during the LGM, haplogroup J sub-lineages arose in the Near Eastern refugia and recolonized Europe following the end of the last glaciation. In particular, J1c is not yet found in any hunter-gatherers, and the oldest individuals belonging to this lineage were found in Iran35 and in Anatolia30 dating to 8000-7700 BCE. It is possible that J1c arrived in Thracia from Anatolia during the early stages of the Neolithic expansion. The expansion of farmers played an important role also in the diffusion of haplogroup T, which has been found in three Thracian samples with the T2b (BM-15 and BM-59A) and T2e (BM-40) sub-lineages. Pala et al.34 particularly suggested that these lineages entered Europe from Anatolia in the Late Glacial period, and have been later diffused around Europe by Neolithic agriculturalists after intermingling with the inhabitants of Southeast Europe. Overall, while haplogroups H, K, J and T arose throughout the Neolithic increasing frequencies in different later communities and present-day European populations, the haplogroup U sub-lineages including U2, U4, U5 and U8 instead mark the genetic pool of European pre-LGM hunter-gatherers36,37,38. |
|
|
|
Post by Admin on Feb 19, 2020 20:54:41 GMT
The mtDNA genetic relationships between Thracians and the other ancient Eurasian populations (Supplementary Table S2) were directly explored through a correspondence analysis (COA, Fig. 3). The first component, which accounted for 28.3% of the total variance, clearly separates all hunter-gatherers from the rest of Neolithic, Bronze Age and Iron-Age population groups. Along the second component (10,6% of variance), the ancient populations appear instead distributed along a cline of genetic variation which extends from the Early Neolithic farmers of Southern Europe and Anatolia to the Late Neolithic/Bronze Age Europeans and Steppe pastoralists, in accordance with the genomic structure of ancient Europe29,30,32,33. From an autosomal genetic perspective, besides showing the clear discontinuity of Paleolithic hunter-gatherers, recent genome-wide aDNA studies, have indeed outlined two opposite genetic components contributing to the European genetic ancestry: i.e. the ancestry of the Early European farmers related to Anatolian farmers and pre-farming Levant populations and, on the other side, the so-called Steppe ancestry eventually spread into Europe and Asia during the Bronze Age migrations of Yamnaya herders. In this scenario, the mtDNA genetic composition of analyzed Thracian population located them in the middle of this cline, clustering closely to the Peloponnese-Neolithic individuals (Peloponnese_N) and the Chalcolithic and Bronze Age populations of the Balkans (Balkans_Chalcolithic, Balkans_BA). This finding seems to support a mitochondrial genetic profile of the Thracians that reflects their geographical position at the gateway of Europe. In a more general perspective, Thracians show a mtDNA genetic composition that is thus intermediate between the western Eurasian and the Mediterranean populations, documenting a prolonged interaction between people of these regions during the Bronze Age. On the other hand, the relatively higher distance with the Bronze Age populations from the Steppe (Steppe_EMBA and Steppe_MLBA), may support the hypothesis that the Thracians largely derived from local people9,10,11 with only a low percentage of the gene flow from the Steppe, at least during the early stages of their cultural development. However, in order to better explore this hypothesis, it is worth emphasizing that the perspective offered here by the analysis of mitochondrial genomes should be integrated by the possibility of testing the results obtained with Y-chromosome and autosomal genome-wide data. At this respect, several studies have indeed pointed out the sex-biased nature of the recent demographic changes and expansions in Eurasia39,40,41,42,43, thus suggesting possible sex-specific patterns of migration. 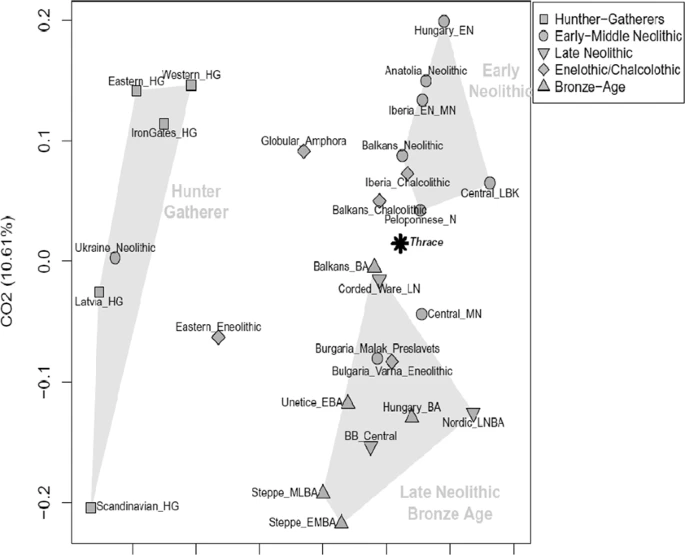 Figure 3 In addition to a temporal frame, in order to explore the spatial pattern of mtDNA genetic variability, the genetic composition of past Thracian population was compared also with that of present-day human groups by means of a spatial Principal Component Analysis (sPCA, Fig. 4). Along the first component (sPC1) the ancient Thracians are closely related with Central-East European populations, while along the second component (sPC2) our samples show higher resemblance with present-day Mediterranean groups. Despite the general lack of statistical support to a clear-cut genetic structure (Gtest: obs = 0.196, P-value = 0.182), as expected due to the well-known higher genetic homogeneity of the mtDNA variability, this pattern reflects the one highlighted by COA analysis on ancient populations. Overall, the mitochondrial genetic structure observed in our sample seems to be mainly a consequence of demographic processes between two macro-areas: West Eurasia and the Mediterranean. This is in agreement with previous studies on modern samples14,15,16 that identify features of both Eastern Europe and Mediterranean area in Bulgarian population. 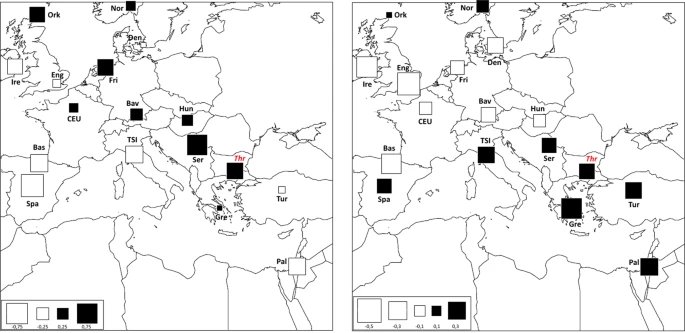 Figure 4 Spatial Principal Component Analysis (sPCA) based on Thracian and modern comparison populations. The first two global components sPC1 (a) and sPC2 (b) are depicted. Positive values are represented by black squares; negative values are represented by white squares; the size of the square is proportional to the absolute value of sPC scores. |
|
|
|
Post by Admin on Feb 19, 2020 22:46:12 GMT
Discussion
In the present study, we reconstructed and analyzed complete mitochondrial genomes from 25 Bronze Age individuals sampled in three Bulgarian necropolises. According to the archaeological records, these cemeteries are associated to the Thracians culture and the chronology, attributed by funerary context, was confirmed by three direct radiocarbon dating placing the remains at II-III millennium BC. These data were used to explore, for the first time, the genetic structure of this ancient population.
We found that the Thracian maternal gene pool is represented essentially by Western Eurasian haplogroups, as expected given the well-known overall mtDNA genetic similarity among all European populations. However, when we compared the complete mitochondrial sequences of Thracians to that of ancient and contemporary Eurasian populations, we observe that their genetic profile reflects their nexus geographical position between east and west.
Several studies demonstrated that Balkan Peninsula has been in different times a crossroad for people moving from and to Europe and beyond16,44. While previous analyses of modern populations demonstrated the impact of such migrations on the genetic makeup of present-day Bulgarians14,15,16, scarce information were available for the ancient (proto-) Bulgarian maternal gene pool and were mainly limited to HVS1 data from the medieval period19. In this study, we provide, for the first time, genetic details of an ancient population, which is particularly relevant from both a chronological and a geographical point of view. In accordance with their geographical location, Thracians show a genetic composition clearly intermediate between East Europe and Mediterranean, that suggests multiple admixture events and population movements occurred across what is now the modern day Bulgaria. Albeit limited to DNA transmitted along the female lines of descent, our genetic data on ancient Thracians provide a direct evidence of how the Balkan region has been a link between East and West Europe since the prehistoric time, and particularly during the Neolithic and post-Neolithic events. In this perspective, future studies will certainly benefit from the analysis of nuclear genome (Y-chromosome and autosomal genetic variation) in order to integrate the observed mtDNA genetic patterns within a more comprehensive overview and for testing the possibility of different sex-biased migrations in the area.
Overall, the ancient mtDNA data presented in this study integrate the existing database and has important implication for understanding the origins of the peopling in this part of Europe and for enlarging the knowledge on the ancient Bronze Age civilizations. How and to what extent ancient Thracian people has contributed to the present-day Bulgarian gene pool remain largely unknown due to the lack of large mitogenomes from contemporary populations from the area, necessary for a phylogenetically and demographically informative comparison.
Scientific Reports volume 9, Article number: 5412 (2019)
|
|

