|
|
Post by Admin on Feb 29, 2020 18:50:37 GMT
The spread of steppe and Iranian-related ancestry in the islands of the western Mediterranean Abstract Steppe-pastoralist-related ancestry reached Central Europe by at least 2500 BC, whereas Iranian farmer-related ancestry was present in Aegean Europe by at least 1900 BC. However, the spread of these ancestries into the western Mediterranean, where they have contributed to many populations that live today, remains poorly understood. Here, we generated genome-wide ancient-DNA data from the Balearic Islands, Sicily and Sardinia, increasing the number of individuals with reported data from 5 to 66. The oldest individual from the Balearic Islands (~2400 BC) carried ancestry from steppe pastoralists that probably derived from west-to-east migration from Iberia, although two later Balearic individuals had less ancestry from steppe pastoralists. In Sicily, steppe pastoralist ancestry arrived by ~2200 BC, in part from Iberia; Iranian-related ancestry arrived by the mid-second millennium BC, contemporary to its previously documented spread to the Aegean; and there was large-scale population replacement after the Bronze Age. In Sardinia, nearly all ancestry derived from the island’s early farmers until the first millennium BC, with the exception of an outlier from the third millennium BC, who had primarily North African ancestry and who—along with an approximately contemporary Iberian—documents widespread Africa-to-Europe gene flow in the Chalcolithic. Major immigration into Sardinia began in the first millennium BC and, at present, no more than 56–62% of Sardinian ancestry is from its first farmers. This value is lower than previous estimates, highlighting that Sardinia, similar to every other region in Europe, has been a stage for major movement and mixtures of people. 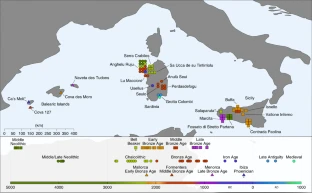 Fig. 1: Geographical origins and temporal distribution of newly reported data
80 Introduction
81 The advent of the European Bronze Age after 3000 BCE was marked by an increase in long-range
82 human mobility. People with ancestry from the Steppe north of the Black and Caspian Seas made a
83 profound demographic impact in central and eastern Europe, mixing with local farmers to
84 contribute up to three quarters of the ancestry of peoples associated with the Corded Ware
85 complex1–3. The expansion of the Beaker complex after around 2400 BCE from the west had a less
86 straightforward correlation to genetic ancestry. In Iberia, most people buried with artifacts of the
87 Beaker complex had little if any Steppe pastoralist-related ancestry (from here on denoted “Steppe
88 ancestry”), but Beaker cultural practices were adopted by people in Central Europe were in part
89 descended from Steppe pastoralists and then spread this material culture along with Steppe
90 ancestry to northwestern Europe4. In Iberia, Steppe ancestry began to appear in outlier individuals
91 by ~2500 BCE4, and became fully mixed into the Iberian population by 2000 BCE5. Meanwhile on
92 Crete in the eastern Mediterranean, there was little if any Steppe ancestry identified in all
93 published samples from the Middle to Late Bronze Age “Minoan” culture (individuals dating to 2400-
94 1700 BCE), although these individuals derived about 15% of their ancestry from groups related to
95 early Iranian farmers (from here on referred to as “Iranian-related ancestry”)6 (Fig. 1).
96 In the islands of the central and western Mediterranean, the Bronze Age, transition has not been
97 investigated with ancient DNA, despite the fact that archaeological evidence reveals that many of
98 the same cultural changes that affected mainland Europe and the eastern Mediterranean also
99 impacted this region7. The first evidence for a permanent human presence in the Balearic Islands is
100 dated to just before the onset of the Bronze Age in this part of Europe, between ~2500-2300 BCE8,9.
101 Early settlers initially relied on animal husbandry and their economy was focused on sheep, goat9,10,
102 and cereal agriculture11, while exploitation of wild marine resources (fish, marine birds, mollusks)
103 was central to subsistence on the small island of Formentera10,12. Around 1200 BCE, the
104 development of the Talaiotic culture in Mallorca and Menorca (the easternmost Balearic Islands)
105 was marked by intensified management of food resources and the appearance of monumental
106 towers, the eponymous talaiots. These structures were similar in style to the Sardinian nuraghi10,13,
107 raising the question of whether there was a cultural connection14, a scenario that would gain
108 plausibility if there was substantial genetic exchange between the two regions. Nuragic Sardinians
109 were also in cultural contact with groups from the eastern Mediterranean15, so an important
110 question is whether they were admixed with either Steppe or Iranian-related ancestry. Meanwhile,
111 the central Mediterranean island of Sicily was affected by the spread of Beaker cultural complex
112 after around 2400 BCE, and by cultural influence from the Aegean in the Late Helladic Period
113 ~1600-1200 BCE (the period of the “Mycenaean” culture)16–18. An unanswered question is whether
114 these events or other cultural changes on the island involved substantial movements of people.
115 We increased the number of individuals from these islands with genome-wide data from 3 to 52,
116 and analyzed the data to address three questions. First, to what extent did movements of people
117 into these islands track the material culture exchanges documented in the archaeological record?
118 Secondly, can we establish the source and minimum dates of arrival of Steppe ancestry in the
119 central and western Mediterranean islands where this ancestry is present in variable proportions
120 today? Thirdly, did Iranian-related ancestry reach the central and western Mediterranean prior to
121 the period of Phoenician and Greek expansion?
|
|
|
|
Post by Admin on Mar 1, 2020 18:35:03 GMT
122 Results
123 Samples and sequencing results
124 We prepared powder from petrous bones and teeth in dedicated ancient DNA clean rooms at
125 University College Dublin, Harvard Medical School, and the University of Florence, extracted DNA
126 using a method designed to retain short molecules19–22, and converted the extracted DNA into
127 double-stranded libraries23. We treated all libraries with Uracil-DNA Glycosylase (UDG) to cleave
128 the analyzed molecules at damaged uracil sites, thereby greatly reducing the rate of cytosine-to
129 thymine errors characteristic of ancient DNA. We enriched ancient DNA libraries for sequences
130 overlapping approximately 1.24 million single nucleotide polymorphisms (SNPs)24,25, and obtained
131 genome-wide data from a total of 49 individuals from the Balearic Islands, Sardinia, and Sicily while
132 increasing the quality of data for a Bell Beaker culture associated individual from Sicily (adding
133 three more libraries to the one previously generated) (Fig. 1, Online Table 1, Supplementary
134 Materials). We established chronology based on archaeological context and by assembling direct
135 radiocarbon dates on bone for 28 of the individuals (direct dates for 26 individuals are reported for
136 the first time here; Online Table 2). We removed from the analysis dataset eight individuals with
137 fewer than 20,000 of the targeted SNPs covered by at least one sequence, and five with evidence of
138 substantial contamination or less than 3% cytosine-to-thymine error in terminal cytosines. We also
139 removed one individual who we detected as a first degree relative (a son) of another (his mother)
140 that gave higher quality genetic data. This left 36 individuals for our modeling (however, since all
141 the data are useful we fully report all individuals, Online Table 1). In the analysis dataset, the
142 median coverage on targeted SNPs on chromosomes 1-22 was 3.20-fold (range 0.02-12.13), and the
143 median number of SNPs covered by at least one sequence was 756709 (range 23600-1038409). All
144 mitochondrial DNA point estimates for match rate to the consensus sequence had 95% confidence
145 intervals with upper bounds from 0.96-1.00, while contamination estimates based on X chromosome
146 variation (meaningful only in males) were all below 1.1% (Online Table 1). All individuals had data
147 from at least one library with cytosine-to-thymine damage in the terminal nucleotides greater than
148 3% (the minimum suggested as a guideline for the plausible authentic DNA23). The qualitative
149 patterns of ancestry in the data were unchanged when we restricted to transversion SNPs which are
150 not affected by characteristic ancient DNA errors (Supplementary Fig. 1).
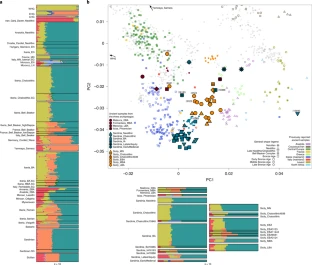 Fig. 2: Overview of the genetic structure.
151 Genetic affinities and population groupings
152 We carried out principal component analysis (PCA) of the ancient individuals merged with
153 previously published ancient DNA1,2,4,6,26–44, projected onto genetic variation among 737 diverse
154 present-day west Eurasians genotyped at ~600,000 SNPs (a subset of the positions on the ~1.24
155 million SNP set)27,39,45–47 (Fig. 2b, Online Table 3). We also performed unsupervised clustering with
156 ADMIXTURE48 (Fig. 2a). The three Balearic Islands individuals in the analysis dataset fall between
157 the European Neolithic and Bronze Age clusters on the PCA, consistent with harboring Steppe
158 ancestry (Fig. 2b), a finding that is also supported by the finding in these individuals of an
159 ADMIXTURE component maximized in Eastern European Hunter Gatherers (EHG) and Yamnaya
160 Steppe Pastoralists. The eight Nuragic Sardinians cluster in PCA and ADMIXTURE with Middle
161 Neolithic Europeans, with the exception of one individual (I10365: 1643-1263 calBCE) that shows a
162 shift towards the Sicilian cluster. The Iron Age Sardinian (~400-200 BCE) and the four Late Antiquity
163 Sardinians (~200-700 CE) deviate toward the Mycenaean cluster, while one of the Late Antiquity
164 Sardinians also deviates toward Central European Bronze Age individuals. The 20 Sicilians cluster in
165 PCA and ADMIXTURE mostly with the European Neolithic individuals, with the exception of two that
166 have more affinity to the Central European Bronze Age individuals (Fig. 2). Relative to the Middle
167 Neolithic Sicilians (Sicily_MN), the main Bronze Age Sicilian cluster (after removing these outliers)
168 deviates in a more subtle way toward eastern groups (either Steppe pastoralists or individuals from
169 the Aegean Bronze Age), a pattern that is also evident in ADMIXTURE.
170 To formally cluster these individuals, we used qpWave2 to test whether each individual in turn was
171 consistent with being from the same group as others from the same time period and region (that is,
172 we tested whether they were consistent with forming a clade at a p<0.010 level) (Supplementary
173 Materials, Fig. 3). In some instances where the qpWave results were ambiguous, we carried out
174 more refined tests to split individuals into analysis groupings (Supplementary Materials).
|
|
|
|
Post by Admin on Mar 2, 2020 6:31:34 GMT
175 In the Balearic Islands, qpWave revealed significant differences between the Early Bronze Age
176 individual Mallorca_EBA and the Late Bronze Age individual Menorca_LBA (p=0.002)
177 (Supplementary Table 1). While qpWave tests comparing the Middle Bronze Age individual
178 Formentera_MBA to the other two individuals were non-significant, the symmetry test statistic
179 f4(Mbuti.DG, Iberia_Chalcolithic; Formentera_MBA, Menorca_LBA) was Z = 2.6 standard errors from
180 zero (exceeding a threshold of |Z|>2, which is approximately p<0.05), implying significantly
181 different ancestry in Formentera_MBA than in Menorca_LBA. In light of this and the different dates
182 and island sources of these three individuals, we treated all three separately for analysis.
183 For the Nuragic Sardinians (Sardinia_Nuragic_BA), only I10365 clearly did not form a clade with
184 others of the same cultural affiliation (Supplementary Tables 2 and 3). Thus, we treated this
185 individual whose radiocarbon date confirms it as contemporaneous with the others as an outlier
186 (Sardinia_Nuragic_BA10365). The Iron Age Sardinian individual formed a clade with some from the
187 Sardinian_LateAntiquity cluster and some ancient Sicilians, but we treated it separately because of
188 its distinctive time period and geographic location. The four Sardinian_LateAntiquity individuals
189 were consistent with forming a clade in qpWave, but one individual separated from the others in
190 PCA (Fig. 2), and also showed distinct signals in admixture modeling, and hence we analyzed it
191 separately as Sardinian_LateAntiquity12221 (Supplementary Table 4).
192 For Sicily, our analysis confirmed the two Early Bronze Age outliers Sicily_EBA11443 and
193 Sicily_EBA8561 evident in PCA and ADMIXTURE (both p<10-12 relative to the main cluster), while
194 identifying a third outlier Sicily_EBA3123 (p=0.004) (Supplementary Tables 5 and 6). One Sicilian
195 Middle Bronze Age individual was not consistent with being a clade with one of the other two, and
196 we treated the three separately in subsequent analysis (Sicily_MBA3124, Sicily_MBA3125, and
197 Sicily_MBA4109) (Supplementary Table 7). All 5 Late Bronze Age individuals were consistent with
198 being a clade at the p>0.01 threshold and we grouped them (Sicily_LBA) (Supplementary Table 8).
199 We used qpAdm2,45 to decompose the ancestry of each analysis grouping into four “distal” sources:
200 Anatolia_Neolithic, Western Hunter-Gatherers (WHG), Iran_Ganj_Dareh_Neolithic and
201 Yamnaya_Samara. We first tested the model with Anatolia_Neolithic and WHG, then added either
202 Iran_Ganj_Dareh_Neolithic or Yamnaya_Samara as a potential third source, and finally combined
203 all ancestry sources for a total of four sources. We quote the most parsimonious model (as
204 measured by the lowest number of ancestry sources) that fits at p>0.05. A unique parsimonious
205 model fit for each analysis grouping (Fig. 4b and Supplementary Table 9 and 10).
 Fig. 3: Pairwise qpWave testing to group individuals.
247 Formal Modeling of Ancestry Changes Over Time in Sardinia
248 We analyzed 13 individuals from Sardinia dated to ~2200 BCE - 700 CE (Fig. 1, Online Table 1).
249 In qpAdm, all eight Bronze Age Nuragic individuals fit as descending from the same two deep
250 ancestral sources (Anatolia_Neolithic and WHG), but mixed in different proportions: 82.5 ± 1.1%
251 Anatolia_Neolithic for the main Sardinia_Nuragic_BA cluster (p=0.265), and 85.4 ± 2.2% for the
252 Sardinia_Nuragic_BA10365 outlier (p=0.064) (Supplementary Table 9). We find no working models
253 when we consider chronological or geographically more proximal sources (e.g. Beaker complex
254 associated individuals from Iberia, France, Czech Republic, Germany; or Chalcolithic Iberians and
255 Neolithic Sicilians), although we do not have access to early Neolithic Sardinians for this analysis.
256 Most Sardinians buried in a Nuragic Bronze Age context possessed uniparental haplogroups found in
257 European hunter-gatherers and early farmers, including Y-haplogroup R1b1a[xR1b1a1a] which is
258 different from the characteristic R1b1a1a2a1a2 spread in association with the Bell Beaker complex4.
259 An exception is individual I10553 (1226-1056 calBCE) who carried Y-haplogroup J2b2a (Online Table
260 1), previously observed in a Croatian Middle Bronze Age individual bearing Steppe ancestry44,
261 suggesting the possibility of genetic input from groups that arrived from the east after the spread
262 of first farmers. This is consistent with the evidence of material culture exchange between
263 Sardinians and mainland Mediterranean groups15, although genome-wide analyses find no significant
264 evidence of Steppe ancestry so the quantitative demographic impact was minimal. qpAdm modeling
265 of the ancestry of the Sardinia_Nuragic_BA10365 outlier with respect to sources potentially more
266 closely related in space and time does infer some ancestry in this individual from an eastern source
267 (either carrying Steppe ancestry or Iranian-related ancestry) that we do not detect by modeling
268 with sources more distant in space and time, consistent with the hypothesis of eastern influence
269 (Supplementary Table 12).
270 We detect definitive evidence of Iranian-related ancestry in an Iron Age Sardinian I10366 (391-209
271 calBCE) with an estimate of 11.9 ± 3.7.% Iran_Ganj_Dareh_Neolithic related ancestry, while
272 rejecting the model with only Anatolian_Neolithic and WHG at p=0.0066 (Supplementary Table 9).
273 The only model that we can fit for this individual using a pair of populations that are closer in time
274 is as a mixture of Iberia_Chalcolithic (11.9 ± 3.2%) and Mycenaean (88.1 ± 3.2%) (p=0.067). This
275 model fits even when including Nuragic Sardinians in the outgroups of the qpAdm analysis, which is
276 consistent with the hypothes is that this individual had little if any ancestry from earlier Sardinians.
277 In the Sardinian_LateAntiquity group (the earliest dating to 256-403 calCE), we detect even higher
278 proportions of Iran_Ganj_Dareh_Neolithic-related ancestry: an estimated 29.6 ± 4.6.% (p=0.000001
279 for rejection of the alternative model that attempts to model its eastern ancestry as entirely
280 Yamnaya-related, Supplementary Table 9). One possibility is the Iranian-related ancestry began to
281 be introduced in the Phoenician period, a scenario that is not only consistent with the historical
282 evidence and our finding of this ancestry type in the Iron Age Sardinian, but is also supported by
283 previously published mitochondrial DNA which has documented haplotypes in ancient Phoenician
284 colonies in modern Sardinians51. In modeling using source populations that are temporally more
285 plausible, this individual is consist with being a clade with both Myceanean (p=0.241) or
286 Ibiza_Phoenician (p=0.145); importantly, both these models works with Nuragic Bronze Age
287 Sardinians included in the outgroups, and so Sardinian_LateAntiquity is consistent with having
288 negligible ancestry from earlier Bronze Age groups to the limits of our resolution (Supplementary
290 Materials). We also model the outlier Sardinia_LateAntiquity12221 as having 33.3 ± 5.5% Yamnaya
300 related while confidently rejecting models with no Steppe ancestry (all p≤0.001) (Supplementary
291 Table 9), providing the earliest clear evidence of Steppe ancestry in Sardinia. However, we do not
292 have sufficient resolution given the limited data from this single sample to determine the
293 geographic source of the Steppe ancestry (Supplementary Table 13).
294 In a dataset of 27 modern Sardinians for whom we have genotyping data at about 600,000 SNPs45,
295 we obtain a fit for a model of 61.4 ± 1.6% Anatolia_Neolithic, 9.5 ± 1.0% WHG, 19.1 ± 1.9%
296 Iran_Ganj_Dareh_Neolithic and 10.0 ± 1.6% Yamnaya_Samara related ancestry and definitively
297 reject models without all four ancestries (all models p<10-6 in Supplementary Table 9). We
298 replicate the finding of Iran_Ganj_Dareh_Neolithic-related ancestry (and not just Steppe ancestry)
299 in a subset of four of the modern Sardinian individuals with whole genome shotgun sequencing data
300 (Supplementary Table 9). Even the four-way model is not comprehensive for modern Sardinians,
301 however, as when we add Late Neolithic North Africans from Morocco to the outgroup set52, we
302 reject the four-way mixture model (p<10-12) (adding the Neolithic Moroccans to the outgroup set
303 does not cause model rejection for any of the ancient samples in our dataset, showing that it may
304 reflect events taking place after the times our individuals lived; Supplementary Table 9). Modeling
305 modern Sardinians with this fifth sources produces a fit with an estimate of 16.1 ± 8.4%
306 Morocco_LN-related ancestry (p=0.235). Our signal of North African-related mixture in Sardinians
307 may reflect the same process that introduced sub-Saharan African ancestry into Sardinians53–55
308 which was argued in 56 to reflect North African-related admixture with an average date of ~630 CE.
309 An important question is how much ancestry modern Sardinians have inherited from people related
310 to those of the Nuragic Bronze Age. We could parsimoniously model our modern Sardinian sample as
311 a 2-way mixture of 13.6 ± 3.4% Sardinia_Nuragic_BA and 86.4 ± 3.4% Sardinia_LateAntiquity12221.
312 It is striking that most of the ancestry in modern Sardinians is inferred in this analysis to come from
313 a Sardinia_LateAntiquity12221-related group, which can itself be modeled as closely related to
314 Mycenaeans or Phoenicians with no evidence of specific shared ancestry with Bronze Age Sardinians.
315 The group of modern Sardinians we are modeling has often been interpreted as an isolated lineage
316 that derives from early Sardinian farmers with little subsequent immigration into the islands. Our
317 finding that a large fraction of this group’s ancestry is consistent with deriving from a group that
318 was present in Sardinia in Late Antiquity and that had no evidence of a contribution from earlier
319 Sardinian groups is therefore surprising (although we caution that this inference is tentative as the
320 amount of data we have for Sardinia_LateAntiquity12221 is limited; Online Table 1). Modern
321 Sardinian populations are geographically highly substructured for example among different valleys
322 and coastal and inland sites.55 Analyses of more geographically diverse modern and ancient
323 Sardinians will provide additional insight into the population turnovers.
|
|
|
|
Post by Admin on Mar 3, 2020 1:20:29 GMT
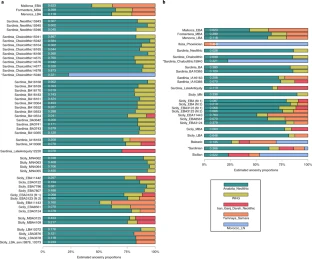 Fig. 4: Distal modelling of ancestry proportions
324 Formal Modeling of the Neolithic to Bronze Age transition in Sicily
325 In the Middle Neolithic, Sicilians harbored ancestry typical of early European farmers, well modeled
326 as a mixture of Anatolia_Neolithic and WHG (Fig. 2, Fig. 4, Supplementary Table 9).
327 Steppe ancestry arrived in Sicily by the Early Bronze Age. While a previously reported Bell Beaker
328 culture-associated individual from Sicily had no evidence of Steppe ancestry4, a result we confirm
329 by more than tripling the number of sequences for this individual who previously had marginal
330 quality data, we find evidence of Steppe ancestry in the Early Bronze Age by ~2200 BCE. In distal
331 qpAdm, the outlier Sicily_EBA11443 is parsimoniously modeled as harboring 40.2 ± 3.5% Steppe
332 ancestry, and the outlier Sicily_EBA8561 is parsimoniously modeled as harboring 23.3 ± 3.5% Steppe
333 ancestry (Fig. 4a, Supplementary Table 9). The main Sicily_EBA cluster also can only be fit with
334 Steppe ancestry albeit at a lower proportion of 9.1 ± 2.3%, and models without Steppe ancestry can
335 be rejected (p=0.001) (Supplementary Table 9). The presence of Steppe ancestry in Early Bronze
336 Age Sicily is also evident in Y chromosome analysis, which reveals that 4 of the 5 Early Bronze Age
337 males had Steppe-associated Y-haplogroup R1b1a1a2a1a2. (Online Table 1). Two of these were Y
338 haplogroup R1b1a1a2a1a2a1 (Z195) which today is largely restricted to Iberia and has been
339 hypothesized to have originated there 2500-2000 BCE57. This evidence of west-to-east gene flow
340 from Iberia is also suggested by qpAdm modeling where the only parsimonious proximate source for
341 the Steppe ancestry we found in the main Sicily_EBA cluster is Iberians (Supplementary Table 14).
342 We detect Iranian-related ancestry in Sicily by the Middle Bronze Age 1800-1500 BCE, consistent
343 with the directional shift of these individuals toward Mycenaeans in PCA (Fig. 2b). Specifically, two
344 of the Middle Bronze Age individuals can only be fit with models that in addition to
345 Anatolia_Neolithic and WHG, include Iran_Ganj_Dareh_Neolithic. The most parsimonious model for
346 Sicily_MBA3125 has 18.0 ± 3.6% Iranian-related ancestry (p=0.032 for rejecting the alternative
347 model of Steppe rather than Iranian-related ancestry), and the most parsimonious model for
348 Sicily_MBA4109 has 14.9 ± 3.9% Iranian-related ancestry (p=0.037 for rejecting the alternative
349 model) (Fig. 4a, Supplementary Table 9). This inference is also supported by qpAdm using sources
350 closer in geography and time that always identify a parsimonious model with Minoan_Lassithi as a
351 source for these two individuals (Supplementary Table 15). We also found evidence of Iranian
352 related ancestry in Sicily in an individual of the Early Bronze Age cluster, I11442, who could only be
353 fit in a 3-way model with Iranian-related ancestry (19.3 ± 3.8% ancestry of this type, p=0.391; the
354 3-way model involving Steppe ancestry fails to a fit (p=0.010)) (Supplementary Table 10). However,
355 this finding should be viewed with caution as qpWave clustered this individual with four other
356 Sicilian Early Bronze Age individuals, so this finding could be an artifact of performing tests on our
357 data beyond what is justified by our groupings. The modern southern Italian Caucasus-related signal
358 identified in 58 is plausibly related to the same Iranian-related spread of ancestry into Sicily that we
359 observe in the Middle Bronze Age (and possibly the Early Bronze Age).
360 For the Late Bronze Age group of individuals, qpAdm documented Steppe-related ancestry,
361 modeling this group as 80.2 ± 1.8% Anatolia_Neolithic, 5.3 ± 1.6% WHG, and 14.5 ± 2.2%
362 Yamnaya_Samara (Fig. 4b, Supplementary Table 9). Our modeling using sources more closely
363 related in space and time also supports Sicily_LBA having Minoan-related ancestry or being derived
364 from local preceding populations or individuals with ancestries similar to those of Sicily_EBA3123
365 (p=0.527), Sicily_MBA3124 (p=0.352), and Sicily_MBA3125 (p=0.095) (Supplementary Table 15).
366 Finally, when we model modern Sicilians, we find that they require not only Steppe and Iranian
367 related ancestries but also North African ancestry, confirming the ample historical and
368 archaeological evidence of major cultural impacts on the island from North Africa after the Bronze
369 Age (Supplementary Materials).
|
|
|
|
Post by Admin on Mar 3, 2020 21:09:14 GMT
 The Mediterranean Sea has been a major route for maritime migrations as well as frequent trade and invasions during prehistory, yet the genetic history of the Mediterranean islands is not well documented despite recent developments in the study of ancient DNA. An international team led by researchers from the University of Vienna, Harvard University and University of Florence, Italy, is filling in the gaps with the largest study to date of the genetic history of ancient populations of Sicily, Sardinia and the Balearic Islands, increasing the number of individuals with reported data from 5 to 66. The results reveal a complex pattern of immigration from Africa, Asia and Europe which varied in direction and its timing for each of these islands. For Sicily, the article reports on a new ancestry during the Middle Bronze Age that chronologically overlaps with the Greek Mycenaean trade network expansion.  Sardinians descend from Neolithic farmers A very different story is unraveled in the case of Sardinia. Despite contacts and trade with other Mediterranean populations, ancient Sardinians retained a mostly local Neolithic ancestry profile until the end of the Bronze Age. However, during the second half of the 3rd millennium BC, one of the studied individuals from Sardinia has a large proportion of North African ancestry. Taken together with previous results of a contemporary central Iberian individual and a later 2nd mill. BC Bronze Age individual from Iberia, it clearly shows prehistoric maritime migrations across the Mediterranean Sea from North Africa to locations in southern Europe, affecting more than 1 percent of individuals reported in the ancient DNA literature from this region and time to date. “Our results show that maritime migrations from North Africa started long before the era of the eastern Mediterranean seafaring civilizations and moreover were occurring in multiple parts of the Mediterranean,” says Ron Pinhasi, a co-senior author of the Department of Evolutionary Anthropology, University of Vienna. During the Iron Age expansion and establishment of Greek and Phoenician colonies in the West Mediterranean islands, the two Sardinian individuals analyzed from that period had little, if any, ancestry from the previous long-established populations. “Surprisingly, our results show that despite these population fluxes and mixtures, modern Sardinians retained between 56-62 percent of ancestry from the first Neolithic farmers that arrived in Europe around 8000 years ago,” says David Caramelli a co-senior author, and Director of Department of Biology at the University of Florence. Migration from the Iberian Peninsula documented “One of the most striking findings is about the arrival of ancestry from the Steppe north of the Black and Caspian Seas in some of the Mediterranean islands. While the ultimate origin of this ancestry was Eastern Europe, in the Mediterranean islands it arrived at least in part from the west, namely from Iberia,” says David Reich, a co-senior author at Harvard University, who is also an investigator of the Howard Hughes Medical Institute and at the Broad Institute of MIT and Harvard. “This was likely the case for the Balearic Islands, in which some early residents probably derived at least part of their ancestry from Iberia,” says first author Daniel Fernandes, of the Department of Evolutionary Anthropology, University of Vienna. Reference: “The spread of steppe and Iranian-related ancestry in the islands of the western Mediterranean” by Daniel M. Fernandes, Alissa Mittnik, Iñigo Olalde, Iosif Lazaridis, Olivia Cheronet, Nadin Rohland, Swapan Mallick, Rebecca Bernardos, Nasreen Broomandkhoshbacht, Jens Carlsson, Brendan J. Culleton, Matthew Ferry, Beatriz Gamarra, Martina Lari, Matthew Mah, Megan Michel, Alessandra Modi, Mario Novak, Jonas Oppenheimer, Kendra A. Sirak, Kristin Stewardson, Kirsten Mandl, Constanze Schattke, Kadir T. Özdoğan, Michaela Lucci, Gabriella Gasperetti, Francesca Candilio, Gianfranca Salis, Stefania Vai, Edgard Camarós, Carla Calò, Giulio Catalano, Marián Cueto, Vincenza Forgia, Marina Lozano, Elisabetta Marini, Margherita Micheletti, Roberto M. Miccichè, Maria R. Palombo, Damià Ramis, Vittoria Schimmenti, Pau Sureda, Luís Teira, Maria Teschler-Nicola, Douglas J. Kennett, Carles Lalueza-Fox, Nick Patterson, Luca Sineo, Alfredo Coppa, David Caramelli, Ron Pinhasi and David Reich, 24 February 2020, Nature Ecology & Evolution. DOI: 10.1038/s41559-020-1102-0 |
|

