Post by Admin on Jul 11, 2019 18:14:07 GMT
The new Denisovan hemimandible from Xiahe, China (1), exhibits a 3-rooted lower molar, providing a direct morphological link between archaic and recent Asian Homo sapiens populations. Although mandibular molars are most commonly 2-rooted in the genus Homo, root number varies from 1 to 3 (2) or more (3). Three-rooted lower molars maintain both mesial and distal roots, with a third accessory root on either the distolingual aspect or lingually between the mesial and distal roots (Fig. 1). The third root is not a simple bifurcation of either the mesial or distal root tips. While the accessory root can be quite small, it is usually about one-third the size of the normal roots (2). In recent humans, the third root usually occurs on the mandibular first molar (referred to as a 3RM1) but may also occur on the lower second and third molars; we refer to these collectively as 3RM (4⇓–6). The 3RM may appear either unilaterally or bilaterally; the single twin study of which we are aware shows bilateral development in both twins, suggesting a genetic underpinning (7). The 3RM entered the clinical literature in 1844 (8), being called radix entomolaris by Bolk (9); it was codified into the Arizona State University Dental Anthropology System in 1991 (2).

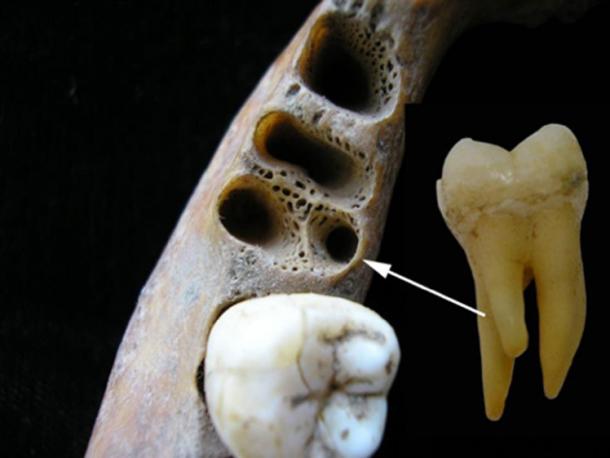
Fig. 1.
The 3-rooted lower molar anomaly. Three-rooted lower first molar alveolar sockets showing distolingual position of accessory root and the 3-rooted lower first molar (lingual view); (Inset) 3-rooted lower second molar of Xiahe Denisovan individual (lingual view). Left and Middle images courtesy of Christine Lee (California State University, Los Angeles, CA). M, mesial; L, lingual; D, distal; B, buccal.
Extensive clinical and bioarchaeological studies confirm the rarity of 3RM outside of Asia and the New World (Dataset S1). In Asian-derived populations, the frequency of the 3RM can exceed 40% (Aleut, Neolithic China), whereas, in non-Asian−derived populations, the frequency ranges from 0 to 3.4%. The rarity of the 3RM in non-Asian H. sapiens is low enough to be explained by mutation alone (2). The high frequencies of 3RM in northeast Asians and Native Americans is a key feature linking Native American origins to Asia (10).

Despite its high frequency in recent Asian-derived populations, the 3RM has not been reported in the earliest H. sapiens from Asia (11), nor have we observed the trait in early H. sapiens from Africa or Homo erectus in Asia.* We note, however, that the lack of radiography of many specimens and the absence of the original Zhoukoudian remains make this conclusion preliminary. Before the recent discoveries, the earliest example of a 3RM came from an H. sapiens mandible from the Philippines (15), perhaps from the site of Tabon, which has fossil-bearing strata dating to 9 ka, 16.5 ka, and as much as 47 ka, and more recent Jar burials (16, 17). The mandible, originally described by Macintosh et al. (18), shows a bilateral 3RM with an accessory root situated lingually between the mesial and distal roots (15). Given a lack of early evidence for 3RM, one explanation for its high frequency in Asia has been a relatively recent acquisition postdating the origin of H. sapiens and occurring well after their dispersal into Eurasia.
Two recently described mandibles suggest a more ancient Asian origin for 3RM, and one that precedes H. sapiens in the region. The newly discovered individual from Xiahe, China—identified as Denisovan through paleoproteomics (1)—possesses a 3-rooted lower second molar (3RM2) (Fig. 1).† This individual is dated to 160 ka. The recently described Penghu 1 mandible from Taiwan (190 to 10 ka) also exhibits a 3RM2 (19). Although the authors suggest the 3RM2 differs from that described by Turner et al. (2), it is clear that the morphology of Penghu 1 falls within the variation described by previous studies (2, 10): The third root appears lingually as an accessory root between the mesial and distal roots. The Penghu mandible retains “archaic” features, including a receding symphysis that lacks a chin, a thick mandibular corpus, and large molar crowns similar in size to Denisovans (20). Like Xiahe, these exceptionally large molars are coupled with agenesis of the third molar (19). For these reasons, Chen et al. (1) suggest that Penghu 1 may also be closely related to Denisovans. Both mandibles show that the 3RM anomaly existed in archaic Asian hominins before H. sapiens in the region.
These 2 recently reported fossils suggest that the 3RM 1) very likely originated in Asia and 2) evolved in a pre-sapiens population. Moreover, until a 3RM is found in more archaic hominins, it should be understood as a morphological trait that was transferred to H. sapiens through gene flow with Denisovans. Gene flow between H. sapiens and Denisovans has been documented, including a mutation (at EPAS1) related to high-altitude adaptation shared by a Siberian Denisovan and modern Tibetans (21). Importantly, Nepal also shows one of the highest occurrences (25%) of 3RM in East Asia (Dataset S1). Like the high-altitude related mutation, which is retained due to positive selection (21), the retention of 3RM at high frequency in Asia may be related to selection for molar retention in populations with heavy masticatory loading (22). Such selection also explains the lower frequency of 3RM in recent populations with higher Denisovan introgression (e.g., Australia/New Guinea) but demonstrably less masticatory robusticity (23). Alternatively, 3RM frequencies may reflect an indirect influence resulting from selection on another trait under high selection, as has been suggested to be the case for incisor crown morphology and EDAR in North and East Asians and in the New World (24). Whatever the cause, we argue that the 3RM anomaly is an example of a morphological character in recent humans that can be clearly traced to this archaic admixture.
The 3RM is an Asian-derived character that we can definitively trace to Denisovans. Thus, we now have very clear evidence that gene flow between archaic groups and H. sapiens resulted in the transfer of identifiable morphological features. There have been a number of Asian H. sapiens fossils described recently that point to admixture with archaic humans as an explanation for the presence of primitive traits [e.g., Dushan (11) and Tianyuan (25)]. If the 3RM was transferred from archaic humans to H. sapiens, other traits may have been as well. Indeed, the presence of “archaic features” in recent Asians that were once used to suggest continuity from Pleistocene Asian H. erectus (26⇓–28) may also have been obtained by introgression from Denisovans.
PNAS first published July 8, 2019
Fig. 1.
The 3-rooted lower molar anomaly. Three-rooted lower first molar alveolar sockets showing distolingual position of accessory root and the 3-rooted lower first molar (lingual view); (Inset) 3-rooted lower second molar of Xiahe Denisovan individual (lingual view). Left and Middle images courtesy of Christine Lee (California State University, Los Angeles, CA). M, mesial; L, lingual; D, distal; B, buccal.
Extensive clinical and bioarchaeological studies confirm the rarity of 3RM outside of Asia and the New World (Dataset S1). In Asian-derived populations, the frequency of the 3RM can exceed 40% (Aleut, Neolithic China), whereas, in non-Asian−derived populations, the frequency ranges from 0 to 3.4%. The rarity of the 3RM in non-Asian H. sapiens is low enough to be explained by mutation alone (2). The high frequencies of 3RM in northeast Asians and Native Americans is a key feature linking Native American origins to Asia (10).
Despite its high frequency in recent Asian-derived populations, the 3RM has not been reported in the earliest H. sapiens from Asia (11), nor have we observed the trait in early H. sapiens from Africa or Homo erectus in Asia.* We note, however, that the lack of radiography of many specimens and the absence of the original Zhoukoudian remains make this conclusion preliminary. Before the recent discoveries, the earliest example of a 3RM came from an H. sapiens mandible from the Philippines (15), perhaps from the site of Tabon, which has fossil-bearing strata dating to 9 ka, 16.5 ka, and as much as 47 ka, and more recent Jar burials (16, 17). The mandible, originally described by Macintosh et al. (18), shows a bilateral 3RM with an accessory root situated lingually between the mesial and distal roots (15). Given a lack of early evidence for 3RM, one explanation for its high frequency in Asia has been a relatively recent acquisition postdating the origin of H. sapiens and occurring well after their dispersal into Eurasia.
Two recently described mandibles suggest a more ancient Asian origin for 3RM, and one that precedes H. sapiens in the region. The newly discovered individual from Xiahe, China—identified as Denisovan through paleoproteomics (1)—possesses a 3-rooted lower second molar (3RM2) (Fig. 1).† This individual is dated to 160 ka. The recently described Penghu 1 mandible from Taiwan (190 to 10 ka) also exhibits a 3RM2 (19). Although the authors suggest the 3RM2 differs from that described by Turner et al. (2), it is clear that the morphology of Penghu 1 falls within the variation described by previous studies (2, 10): The third root appears lingually as an accessory root between the mesial and distal roots. The Penghu mandible retains “archaic” features, including a receding symphysis that lacks a chin, a thick mandibular corpus, and large molar crowns similar in size to Denisovans (20). Like Xiahe, these exceptionally large molars are coupled with agenesis of the third molar (19). For these reasons, Chen et al. (1) suggest that Penghu 1 may also be closely related to Denisovans. Both mandibles show that the 3RM anomaly existed in archaic Asian hominins before H. sapiens in the region.
These 2 recently reported fossils suggest that the 3RM 1) very likely originated in Asia and 2) evolved in a pre-sapiens population. Moreover, until a 3RM is found in more archaic hominins, it should be understood as a morphological trait that was transferred to H. sapiens through gene flow with Denisovans. Gene flow between H. sapiens and Denisovans has been documented, including a mutation (at EPAS1) related to high-altitude adaptation shared by a Siberian Denisovan and modern Tibetans (21). Importantly, Nepal also shows one of the highest occurrences (25%) of 3RM in East Asia (Dataset S1). Like the high-altitude related mutation, which is retained due to positive selection (21), the retention of 3RM at high frequency in Asia may be related to selection for molar retention in populations with heavy masticatory loading (22). Such selection also explains the lower frequency of 3RM in recent populations with higher Denisovan introgression (e.g., Australia/New Guinea) but demonstrably less masticatory robusticity (23). Alternatively, 3RM frequencies may reflect an indirect influence resulting from selection on another trait under high selection, as has been suggested to be the case for incisor crown morphology and EDAR in North and East Asians and in the New World (24). Whatever the cause, we argue that the 3RM anomaly is an example of a morphological character in recent humans that can be clearly traced to this archaic admixture.
The 3RM is an Asian-derived character that we can definitively trace to Denisovans. Thus, we now have very clear evidence that gene flow between archaic groups and H. sapiens resulted in the transfer of identifiable morphological features. There have been a number of Asian H. sapiens fossils described recently that point to admixture with archaic humans as an explanation for the presence of primitive traits [e.g., Dushan (11) and Tianyuan (25)]. If the 3RM was transferred from archaic humans to H. sapiens, other traits may have been as well. Indeed, the presence of “archaic features” in recent Asians that were once used to suggest continuity from Pleistocene Asian H. erectus (26⇓–28) may also have been obtained by introgression from Denisovans.
PNAS first published July 8, 2019