

Post by Admin on Aug 31, 2023 19:12:55 GMT
Fig. 3: Genetic affinities of Stone Age northwestern Africans and schematic summary of the population history of the Maghreb.
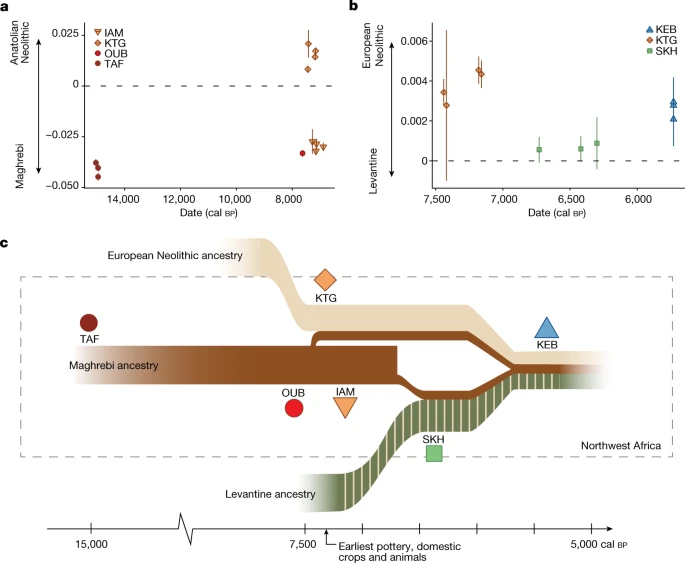
a, Genetic affinity of analysed Stone Age northwestern African individuals, polarized between Neolithic Anatolia and Maghrebi ancestry, using the f4 test of the form f4(Anatolia Neolithic, TAF011; Stone Age northwestern African individuals, Mbuti). b, Genetic affinity of Stone Age northwestern African individuals, polarized between Early Neolithic Iberian and Levantine ancestry using f4 test of the form f4(Iberia Early Neolithic, Levant Chalcolithic; Neolithic northwestern African individuals, Mbuti). a,b, Each symbol represents a single individual f4 value. Error bars indicate ±2 s.e., computed with a block jack-knife approach (5 Mb blocks weighted by the number of SNPs). c, Summary of inferred population history of the Stone Age Maghreb.
Iberian Early Neolithic (both as a whole and regionally) was found to be the best source population for the European ancestry in KTG, followed by Sicily Stentinello Early Neolithic (Supplementary Data 9). This is consistent with low levels of genetic differentiation in Cardial Ware-associated groups along the European shores of the Mediterranean Sea41, confirmed by direct radiocarbon dates showing that Impressed Ware farmers expanded rapidly across the western Mediterranean3,19,42.
It has been debated whether European farmers crossed from Iberia to Morocco2,3 or whether earlier crossings of the Mediterranean would have happened, through the Sicilian–Tunisian Strait followed by a Maghrebi route of expansion4,43. Direct comparisons of Early Neolithic farmers from Sicily and Iberia as ancestors of KTG farmers provide stronger evidence for an Iberian Neolithic origin (Supplementary Data 9 and Supplementary Information 8), but we cannot exclude some contribution from Sicilian farmers. Genetic data are consistent with the most parsimonious explanation for archaeological evidence on the Neolithic transition in northwestern Africa: the crossing from southern Iberia by Iberian Neolithic farmers2,23. The close geographical proximity between southern Iberia and the Tangitana Peninsula adds strength to this observation whereas the lack of reliable archaeological evidence of early domestic elements in relevant sites along the eastern Maghreb and Tunis, including sites with pottery and obsidian from Pantelleria Island, undermines the Sicily–Tunis crossing hypothesis3. Interestingly, gene flow from North Africa was found only in Mediterranean European individuals much later, from around 4,500 years ago31,44.
Different individuals from KTG date to slightly different time periods. We find a twofold larger proportion of Maghrebi ancestry in earlier KTG individuals (roughly 25%, ktg001 and ktg005, approximately 7,429–7,267 cal BP) than in later ones (about 13%, ktg004 and ktg006, around 7,247–6,945 cal BP) (Fig. 1d). This coincides with an increase in European Neolithic ancestry, shown by the significantly negative result for f4(KTG earlier, KTG later, Iberia Early Neolithic, Mbuti; z-score = −5.01). Approximately one quarter of Maghrebi ancestry in early KTG suggests that they represent at least the second generation of interbreeding between the groups. We estimated the time of admixture using two approaches based on ancestry covariance patterns and linkage disequilibrium decay, using Iberia or Sicily Early Neolithic and TAF as admixture sources. Both methods date the contact within the last six to 13 generations (Supplementary Information 8), suggesting that mixing between groups occurred for a few hundred years, which is consistent with analysis of pottery style that points to the first contact at 7,500–7,400 cal BP (ref. 23).
Kaf Taht el-Ghar farmers had slightly lower genetic diversity levels and greater RoH than most Early Neolithic European populations (Fig. 2a,b and Supplementary Fig. 9). The Maghrebi ancestry carried by KTG people shows markedly lower diversity and more extensive RoH, and is probably the cause of the reduction in overall diversity. Archaeological evidence suggests that Early Neolithic farming was restricted to enclaves in westernmost Maghreb, possibly due to climatic constraints to the south4,22. This could have limited the potential of these groups to recover from an initial founder effect.
Overall, the genetic patterns of local interaction between different groups in northwestern Africa are comparable to those found in Europe: farmers assimilated local foragers’ ancestry in a unidirectional admixture process. Cases of hunter-gatherer communities adopting certain elements of the Neolithic have been described in Europe11,14,45. However, the northwestern Africa Neolithization process involved the notable survival of genetically unadmixed local populations (represented by IAM), despite coexisting for at least 300 years with foreign farming communities (KTG), and still adopted several elements of the Neolithic ways of living from them. Whereas the archaeological findings in IAM and KTG point to the exchange of ideas between groups and support an acculturation process of foraging communities1,4, our genetic data show that the exchange of genes was unidirectional.
a, Genetic affinity of analysed Stone Age northwestern African individuals, polarized between Neolithic Anatolia and Maghrebi ancestry, using the f4 test of the form f4(Anatolia Neolithic, TAF011; Stone Age northwestern African individuals, Mbuti). b, Genetic affinity of Stone Age northwestern African individuals, polarized between Early Neolithic Iberian and Levantine ancestry using f4 test of the form f4(Iberia Early Neolithic, Levant Chalcolithic; Neolithic northwestern African individuals, Mbuti). a,b, Each symbol represents a single individual f4 value. Error bars indicate ±2 s.e., computed with a block jack-knife approach (5 Mb blocks weighted by the number of SNPs). c, Summary of inferred population history of the Stone Age Maghreb.
Iberian Early Neolithic (both as a whole and regionally) was found to be the best source population for the European ancestry in KTG, followed by Sicily Stentinello Early Neolithic (Supplementary Data 9). This is consistent with low levels of genetic differentiation in Cardial Ware-associated groups along the European shores of the Mediterranean Sea41, confirmed by direct radiocarbon dates showing that Impressed Ware farmers expanded rapidly across the western Mediterranean3,19,42.
It has been debated whether European farmers crossed from Iberia to Morocco2,3 or whether earlier crossings of the Mediterranean would have happened, through the Sicilian–Tunisian Strait followed by a Maghrebi route of expansion4,43. Direct comparisons of Early Neolithic farmers from Sicily and Iberia as ancestors of KTG farmers provide stronger evidence for an Iberian Neolithic origin (Supplementary Data 9 and Supplementary Information 8), but we cannot exclude some contribution from Sicilian farmers. Genetic data are consistent with the most parsimonious explanation for archaeological evidence on the Neolithic transition in northwestern Africa: the crossing from southern Iberia by Iberian Neolithic farmers2,23. The close geographical proximity between southern Iberia and the Tangitana Peninsula adds strength to this observation whereas the lack of reliable archaeological evidence of early domestic elements in relevant sites along the eastern Maghreb and Tunis, including sites with pottery and obsidian from Pantelleria Island, undermines the Sicily–Tunis crossing hypothesis3. Interestingly, gene flow from North Africa was found only in Mediterranean European individuals much later, from around 4,500 years ago31,44.
Different individuals from KTG date to slightly different time periods. We find a twofold larger proportion of Maghrebi ancestry in earlier KTG individuals (roughly 25%, ktg001 and ktg005, approximately 7,429–7,267 cal BP) than in later ones (about 13%, ktg004 and ktg006, around 7,247–6,945 cal BP) (Fig. 1d). This coincides with an increase in European Neolithic ancestry, shown by the significantly negative result for f4(KTG earlier, KTG later, Iberia Early Neolithic, Mbuti; z-score = −5.01). Approximately one quarter of Maghrebi ancestry in early KTG suggests that they represent at least the second generation of interbreeding between the groups. We estimated the time of admixture using two approaches based on ancestry covariance patterns and linkage disequilibrium decay, using Iberia or Sicily Early Neolithic and TAF as admixture sources. Both methods date the contact within the last six to 13 generations (Supplementary Information 8), suggesting that mixing between groups occurred for a few hundred years, which is consistent with analysis of pottery style that points to the first contact at 7,500–7,400 cal BP (ref. 23).
Kaf Taht el-Ghar farmers had slightly lower genetic diversity levels and greater RoH than most Early Neolithic European populations (Fig. 2a,b and Supplementary Fig. 9). The Maghrebi ancestry carried by KTG people shows markedly lower diversity and more extensive RoH, and is probably the cause of the reduction in overall diversity. Archaeological evidence suggests that Early Neolithic farming was restricted to enclaves in westernmost Maghreb, possibly due to climatic constraints to the south4,22. This could have limited the potential of these groups to recover from an initial founder effect.
Overall, the genetic patterns of local interaction between different groups in northwestern Africa are comparable to those found in Europe: farmers assimilated local foragers’ ancestry in a unidirectional admixture process. Cases of hunter-gatherer communities adopting certain elements of the Neolithic have been described in Europe11,14,45. However, the northwestern Africa Neolithization process involved the notable survival of genetically unadmixed local populations (represented by IAM), despite coexisting for at least 300 years with foreign farming communities (KTG), and still adopted several elements of the Neolithic ways of living from them. Whereas the archaeological findings in IAM and KTG point to the exchange of ideas between groups and support an acculturation process of foraging communities1,4, our genetic data show that the exchange of genes was unidirectional.