Post by Admin on May 15, 2020 4:36:26 GMT
Conclusions
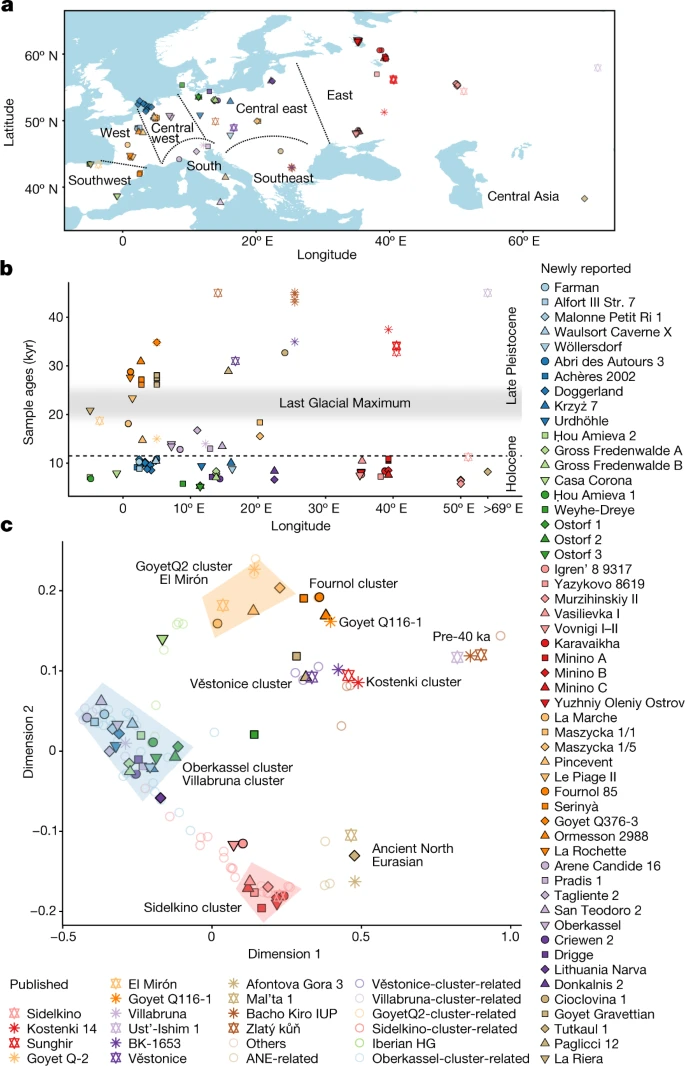
We have shown that the population history of pre-Neolithic Europe was complex in several respects. First, at least some of the initial modern humans to appear in Europe, exemplified by Ust’-Ishim and Oase1, failed to contribute appreciably to the current European gene pool. Only from around 37,000 years ago do all the European individuals analyzed share ancestry with present-day Europeans3. Second, from the time of Kostenki14 about 37,000 years ago until the time of the Villabruna Cluster about 14,000 years ago, all individuals seem to derive from a single ancestral population with no evidence of substantial genetic influx from elsewhere. It is interesting that during this time, the Mal’ta Cluster is not represented in any of the individuals we sampled from Europe. Thus, while individuals assigned to the Gravettian cultural complex in Europe are associated with the Vestonice Cluster, there is no genetic connection between them and the Mal’ta1 individual in Siberia despite the fact that Venus figurines are associated with both. This suggests that if this similarity is not a coincidence32, it reflects diffusion of ideas rather than movements of people. Third, we find that GoyetQ116-1 derives from a different deep branch of the European founder population than the Vestonice Cluster which became predominant in many places in Europe between 34,000 and 26,000 years ago including at Goyet Cave. GoyetQ116-1 is chronologically associated with the Aurignacian cultural complex. Thus, the subsequent spread of the Vestonice Cluster, which is associated with the Gravettian cultural complex, shows that the spread of the latter culture was mediated at least in part by population movements. Fourth, the population represented by GoyetQ116-1 did not disappear, as its descendants became widespread again after ~19,000 years ago in the El Mirón Cluster when we detect them in Iberia. The El Mirón Cluster is associated with the Magdalenian culture and may represent a post-ice age expansion from southwestern European refugia33. Fifth, beginning with the Villabruna Cluster at least ~14,000 years ago, all European individuals analyzed show an affinity to the Near East. This correlates in time to the Bølling-Allerød interstadial, the first significant warming period after the Ice Age34. Archaeologically, it correlates with cultural transitions within the Epigravettian in Southern Europe35 and the Magdalenian-to-Azilian transition in Western Europe36. Thus, the appearance of the Villabruna Cluster may reflect migrations or population shifts within Europe at the end of the Ice Age, an observation that is also consistent with the evidence of turnover of mitochondrial DNA sequences at this time26,37. One scenario that could explain these patterns is a population expansion from southeastern European or west Asian refugia after the Ice Age, drawing together the genetic ancestry of Europe and the Near East. Sixth, within the Villabruna Cluster, some, but not all, individuals have affinity to East Asians. An important direction for future work is to generate similar ancient DNA data from southeastern Europe and the Near East to arrive at a more complete picture of the Upper Paleolithic population history of western Eurasia38.
Extended Data
Extended Data Figure 1

A decrease in Neanderthal ancestry in the last 45,000 years
This is similar to Figure 2, except we use ancestry estimates from rates of alleles matching to Neanderthal rather than f4-ratios, as described in Supplementary Information section 3). The least squares fit excludes Oase1 (as an outlier with recent Neanderthal ancestry) and Europeans (known to have reduce Neanderthal ancestry). The regression slope is significantly negative (P=0.00004, Extended Data Table 3).
Extended Data Fig. 2

Heat matrix of pairwise f3(X, Y; Mbuti) for selected ancient samples
We analyze only samples with at least 30,000 SNPs covered at least once, which pass our quality control.

Extended Data Fig. 3
Studying how the relatedness of non-European populations to pairs of European hunter-gatherers changes over time
We examine statistics of the form D(W, X; Y, Mbuti), with the Z-score given on the y-axis, where W is an early European hunter-gatherer, X is another European hunter-gatherer (in chronological order on the x-axis), and Y is a non-European population (see legend). A: W=Kostenki14. B: W=GoyetQ116-1. C: W=Vestonice16. D: W=ElMiron. |Z|>3 scores are considered statistically significant (horizontal line). The similar Figure 4b gives absolute D-statistic values rather than Z-scores (for W=Kostenki14) and uses pooled regions rather than individual populations Y.
We have shown that the population history of pre-Neolithic Europe was complex in several respects. First, at least some of the initial modern humans to appear in Europe, exemplified by Ust’-Ishim and Oase1, failed to contribute appreciably to the current European gene pool. Only from around 37,000 years ago do all the European individuals analyzed share ancestry with present-day Europeans3. Second, from the time of Kostenki14 about 37,000 years ago until the time of the Villabruna Cluster about 14,000 years ago, all individuals seem to derive from a single ancestral population with no evidence of substantial genetic influx from elsewhere. It is interesting that during this time, the Mal’ta Cluster is not represented in any of the individuals we sampled from Europe. Thus, while individuals assigned to the Gravettian cultural complex in Europe are associated with the Vestonice Cluster, there is no genetic connection between them and the Mal’ta1 individual in Siberia despite the fact that Venus figurines are associated with both. This suggests that if this similarity is not a coincidence32, it reflects diffusion of ideas rather than movements of people. Third, we find that GoyetQ116-1 derives from a different deep branch of the European founder population than the Vestonice Cluster which became predominant in many places in Europe between 34,000 and 26,000 years ago including at Goyet Cave. GoyetQ116-1 is chronologically associated with the Aurignacian cultural complex. Thus, the subsequent spread of the Vestonice Cluster, which is associated with the Gravettian cultural complex, shows that the spread of the latter culture was mediated at least in part by population movements. Fourth, the population represented by GoyetQ116-1 did not disappear, as its descendants became widespread again after ~19,000 years ago in the El Mirón Cluster when we detect them in Iberia. The El Mirón Cluster is associated with the Magdalenian culture and may represent a post-ice age expansion from southwestern European refugia33. Fifth, beginning with the Villabruna Cluster at least ~14,000 years ago, all European individuals analyzed show an affinity to the Near East. This correlates in time to the Bølling-Allerød interstadial, the first significant warming period after the Ice Age34. Archaeologically, it correlates with cultural transitions within the Epigravettian in Southern Europe35 and the Magdalenian-to-Azilian transition in Western Europe36. Thus, the appearance of the Villabruna Cluster may reflect migrations or population shifts within Europe at the end of the Ice Age, an observation that is also consistent with the evidence of turnover of mitochondrial DNA sequences at this time26,37. One scenario that could explain these patterns is a population expansion from southeastern European or west Asian refugia after the Ice Age, drawing together the genetic ancestry of Europe and the Near East. Sixth, within the Villabruna Cluster, some, but not all, individuals have affinity to East Asians. An important direction for future work is to generate similar ancient DNA data from southeastern Europe and the Near East to arrive at a more complete picture of the Upper Paleolithic population history of western Eurasia38.
Extended Data
Extended Data Figure 1
A decrease in Neanderthal ancestry in the last 45,000 years
This is similar to Figure 2, except we use ancestry estimates from rates of alleles matching to Neanderthal rather than f4-ratios, as described in Supplementary Information section 3). The least squares fit excludes Oase1 (as an outlier with recent Neanderthal ancestry) and Europeans (known to have reduce Neanderthal ancestry). The regression slope is significantly negative (P=0.00004, Extended Data Table 3).
Extended Data Fig. 2
Heat matrix of pairwise f3(X, Y; Mbuti) for selected ancient samples
We analyze only samples with at least 30,000 SNPs covered at least once, which pass our quality control.
Extended Data Fig. 3
Studying how the relatedness of non-European populations to pairs of European hunter-gatherers changes over time
We examine statistics of the form D(W, X; Y, Mbuti), with the Z-score given on the y-axis, where W is an early European hunter-gatherer, X is another European hunter-gatherer (in chronological order on the x-axis), and Y is a non-European population (see legend). A: W=Kostenki14. B: W=GoyetQ116-1. C: W=Vestonice16. D: W=ElMiron. |Z|>3 scores are considered statistically significant (horizontal line). The similar Figure 4b gives absolute D-statistic values rather than Z-scores (for W=Kostenki14) and uses pooled regions rather than individual populations Y.