|
|
Post by Admin on Sept 12, 2016 21:08:34 GMT
 The sequencing of entire human mitochondrial DNAs belonging to haplogroup U reveals that this clade arose shortly after the “out of Africa” exit and rapidly radiated into numerous regionally distinct subclades. Intriguingly, the Saami of Scandinavia and the Berbers of North Africa were found to share an extremely young branch, aged merely ∼9,000 years. This unexpected finding not only confirms that the Franco-Cantabrian refuge area of southwestern Europe was the source of late-glacial expansions of hunter-gatherers that repopulated northern Europe after the Last Glacial Maximum but also reveals a direct maternal link between those European hunter-gatherer populations and the Berbers. Because of maternal transmission and lack of recombination, the sequence differentiation of human mtDNA has been generated by only the sequential accumulation of new mutations along radiating maternal lineages. Over the course of time, this process of molecular divergence has given rise to monophyletic units that are called “haplogroups.” Because this process of molecular differentiation occurred mainly during and after the process of human colonization of and diffusion into the different continents and regions, haplogroups and subhaplogroups tend to be restricted to specific geographic areas and population groups (Wallace 1995; Achilli et al. 2004).  Only the founders of the sister superhaplogroups M and N (which includes haplogroup R) (Quintana-Murci et al. 1999) participated in the “out of Africa” exit (Cann et al. 1987; Stringer and Andrews 1988; Cavalli-Sforza et al. 1994; Underhill et al. 2000; Forster et al. 2001) and were successful in colonizing the rest of the Old World. Superhaplogroup N is globally distributed outside Africa, encompassing virtually all of the western Eurasian mtDNA variation, and embraces haplogroup U, nested in haplogroup R. Haplogroup U has an extremely broad geographic distribution that ranges from Europe and North Africa to India and Central Asia and has a very high overall frequency (15%–30%) (Richards et al. 2000; Kivisild et al. 2003; Quintana-Murci et al. 2004) To investigate this striking similarity, the portion of the tree encompassing these two mtDNAs was enriched by sequencing 11 additional mtDNA sequences (22–26, 28, 30, 32, 34, 36, and 37 in fig. 1) bearing the control-region motif 16270-150, a motif found generally at low frequencies (<2%) in Berber populations and in other African groups (such as the Fulbe) known to have intermingled with Berbers (Rosa et al. 2004). The motif also shows similarly low frequencies in virtually all European populations, except the Saami of northern Scandinavia, in which it reaches ∼48% (Tambets et al 2004). Because virtually all Saami mtDNAs with 16270-150 harbor the transitions at nts 16189 and 16144 seen in the Yakut mtDNA, three Saami mtDNAs were included among the additional samples.  The average sequence divergence (± SE, computed as per Saillard et al. [2000]) of the 39 coding-region sequences from the root of haplogroup U was 11.4 ± 1.3 substitutions (disregarding indels and pathological mutations), a value which corresponds—according to the mutation-rate estimate of Mishmar et al. (2003)—to a coalescence time estimate of 58.8 ± 6.8 thousand years (ky) for the entire haplogroup U. This value agrees well with the corresponding estimate of 61.6 ± 12.5 ky, based on the hypervariable segment I (HVS-I) mutation rate (Forster et al. 1996), for these 39 mtDNAs. An age of ∼60 ky indicates that haplogroup U arose very soon after the “out of Africa” exit. As for U5, its sequence divergence was 8.1 ± 1.8 substitutions, corresponding to 41.4 ± 9.2 ky, a time estimate in full agreement with its proposed proto-European origin (Richards et al. 2000). It is striking that the sequence divergence of U5b1b, the subclade encompassing mtDNAs from the Saami, Yakut, Berbers, and Fulbe, was 1.7 ± 0.5 substitutions, thus corresponding to only 8.6 ± 2.4 ky. Such a recent common ancestry of maternal lineages found in populations living as far as 9,000 miles apart and whose anthropological affinities are not at all obvious is, to say the least, unexpected. Can we provide a reasonable explanation? The recent molecular dissection of other mtDNA haplogroups reveals some clues. H1 and H3, two frequent subhaplogroups of H, display frequency peaks centered in Iberia and surrounding populations, including the Berbers of Morocco, and coalescence ages of ∼11 ky (Achilli et al. 2004). Furthermore, their frequency patterns and ages resemble those reported for haplogroup V (Torroni et al. 2001a)—which, similar to U5b1b, is extremely common only in the Saami (together, U5b1b and V encompass almost 90% of the Saami mtDNAs) (Torroni et al. 1996; Tambets et al. 2004). Thus, although these previous studies have highlighted the role of the Franco-Cantabrian refuge area as a major source of the hunter-gatherer populations that gradually repopulated much of central and northern Europe when climatic conditions began to improve ∼15 ky ago, the identification of U5b1b now unequivocally links the maternal gene pool of the ancestral Berbers to the same refuge area and indicates that European hunter-gatherers also moved toward the south and, by crossing the Strait of Gibraltar, contributed their U5b1b, H1, H3, and V mtDNAs to modern North Africans. AJHG Volume 76, Issue 5, May 2005, Pages 883–886 |
|
|
|
Post by Admin on Dec 4, 2018 18:01:53 GMT
 Researchers have found evidence for how humans first spread from Siberia to Finland and Northwest Russia thousands of years ago. Published in the journal Nature Communications, scientists from the Max-Planck-Institute for the Science of Human History and the University of Helsinki studied DNA from 3,500-year-old bones and teeth on the Kola Peninsula in Russia, and a 1,500-year-old water burial in Finland, the first ancient DNA evidence analyzed in Finland. They found that Siberian ancestry from the Kola Peninsula in Northwest Russia spread to populations living in Finland. They also found that the modern-day Saami people, who are indigenous to the northern part of Scandinavia today, once lived further south in Finland.  Comparing these to modern-day Saami, Finnish, and other Uralic language speakers, they found that the oldest samples were the ones with the highest proportion of Siberian ancestry. Among modern European humans, the Saami have the highest proportion, while the Nganasan people from northern Siberia have the largest in the world. The findings show that ancient Finnish people likely lived a mobile and nomadic life, trading and moving over a large range. And the population in Levänluhta was found to be more closely related to modern-day Saami people than to the non-Saami Finnish population.  "This is the first exploration of ancient DNA from Finland and the results are very interesting," Stephan Schiffels from the Max Planck Institute, co-senior author on the study, said in the statement. "However more ancient DNA studies from the area will be necessary to better understand whether the patterns we've seen are representative of Finland as a whole." "Our results show that there was a strong genetic connection between ancient Finnish and ancient Siberian populations," Thiseas Lamnidis from the Max Planck Institute, co-first author of the study said in a statement. Lamnidis suggested that “ancient populations from Siberia may have also shared a subsistence strategy, languages and/or cultural behaviors with Bronze Age and Iron Age Finns, despite the large geographical distance."  |
|
|
|
Post by Admin on Dec 5, 2018 18:08:35 GMT
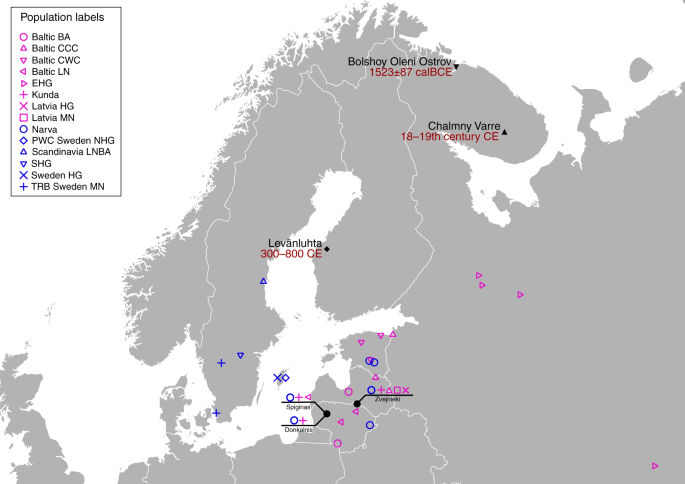 Fig. 1 Sample information and archaeological background The ancient individuals analysed in this study come from three time periods (Table 1, Fig. 1, Supplementary Note 1). The six early Metal Age individuals were obtained from an archaeological site at Bolshoy Oleni Ostrov in the Murmansk Region on the Kola Peninsula (Bolshoy from here on). The site has been radiocarbon dated to 1610-1436 calBCE (see Supplementary Note 1) and the mitochondrial DNA HVR-I haplotypes from these six individuals have been previously reported24. Today, the region is inhabited by Saami. Seven individuals stem from excavations in Levänluhta, a lake burial in Isokyrö, Finland. Artefacts from the site have been dated to the Finnish Iron Age (300–800 CE)25,26. Today, the inhabitants of the area speak Finnish and Swedish. Two individuals were obtained from the 18–19th century Saami cemetery of Chalmny Varre on the Russian Kola Peninsula. The cemetery and the surrounding area were abandoned in the 1960s because of planned industrial constructions, and later became the subject of archaeological excavations. In addition, we sequenced the whole genome of a modern Saami individual to 17.5-fold coverage, for whom genotyping data has previously been published1. Eastern genetic affinities in Northern Europe To investigate the genetic affinities of the sampled individuals, we projected them onto principal components (PC) computed from 1320 modern European and Asian individuals (Fig. 2a, Supplementary Figures 3a, b for a version focusing on West Eurasia). As expected, PC1 separates East Asian from West Eurasian populations. Within each continental group, genetic variability is spread across PC2: The East Asian genetic cline contains populations between the Siberian Nganasan (Uralic speakers) and Yukagirs at one end, and the Ami and Atayal from Taiwan at the other end. The West Eurasian cline along PC2 spans from the Bedouins on the Arabian Peninsula to north-eastern Europeans including Lithuanians, Norwegians and Finns. Between these two main Eurasian clines exist multiple clines, spanning between West and East Eurasians. These clines are likely the result of admixture events and population movements between East and West Eurasia. Most relevant to the populations analysed here is the admixture cline between north-eastern Europe and the North Siberian Nganasan, including mostly Uralic-speaking populations in our dataset (marked in light purple in Fig. 2a). 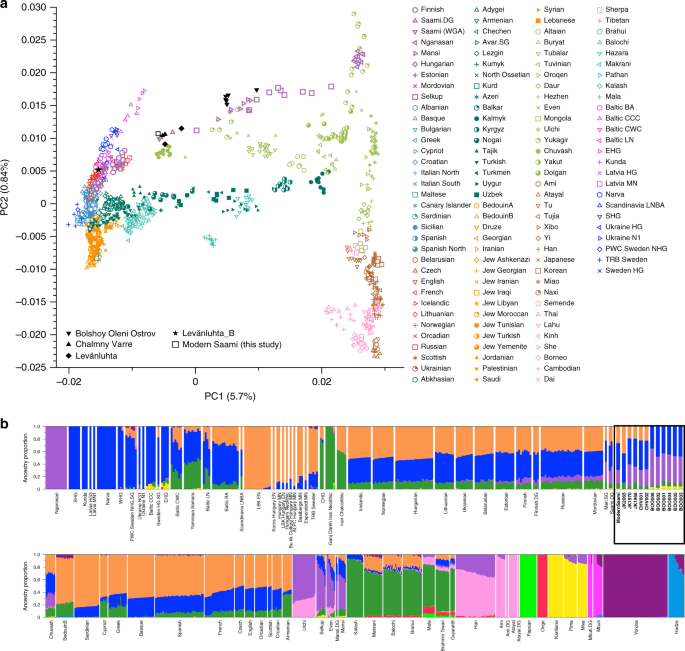 Fig. 2 Ten of the eleven ancient individuals from this study fall on this Uralic cline, with the exception of one individual from Levänluhta (ID JK2065, here named Levänluhta_B), who instead is projected closer to modern Lithuanian, Norwegian and Icelandic populations. Specifically, two Levänluhta individuals and the two historical Saami from Russia are projected very close to the two previously published modern Saami (Saami.DG)32 and the new Saami shotgun genome generated in this study (as well as the previously published genome of the same individual, here labelled Saami (WGA)1), suggesting genetic continuity in the north from the Iron Age to modern-day Saami populations. In contrast, the six ancient individuals from Bolshoy are projected much further towards East Asian populations, and fall to an intermediate position along the Uralic cline and close to modern-day Mansi. Unsupervised genetic clustering analysis as implemented in the ADMIXTURE28 program suggests a similar profile to the PCA: north-eastern European populations harbour a Siberian genetic component (light purple) maximized in the Nganasan (Fig. 2b, see Supplementary Figure 4a for results over multiple K values). The hunter-gatherer genetic ancestry in Europeans (blue) is maximized in European Upper Palaeolithic and Mesolithic hunter-gatherers, including the 8000-year-old Western European hunter-gatherers from Hungary and Spain (WHG), the 8000-year-old Scandinavian hunter-gatherers from Motala (SHG) and the Narva and Kunda individuals from the Baltics. An ancestry component associated with Europe’s first farmers (orange) is maximized in Early Neolithic Europeans associated with the LBK (from German: Linearbandkeramik). The steppe ancestry component within modern Europeans (green), which is associated with the Yamnaya population, is maximised in ancient Iranian populations and to a lesser extent Caucasus hunter-gatherers (CHG). This ancestry component is also present in modern Armenians from the Caucasus, Bedouins from the Arabian Peninsula and South Asian populations. Within modern Europeans, the Siberian genetic component (light purple) is maximized in the Mari and Saami, and can also be seen in similar proportions in the historical Saami from Chalmny Varre and in two of the Levänluhta individuals. The third Levänluhta individual (Levänluhta_B), however, lacks this Siberian component. The six ancient individuals from Bolshoy show substantially higher proportions of the Siberian component: it comprises about half of their ancestry (42.3–58.2%), whereas the older Mesolithic individuals from Motala (SHG) do not possess it at all. The Native-American-related ancestry seen in the EHG and Bolshoy corresponds to a previously reported affinity towards Ancient North Eurasians (ANE)2,33 contributing genes to both Native Americans and West Eurasians. ANE ancestry also comprises part of the ancestry of Nganasans2. 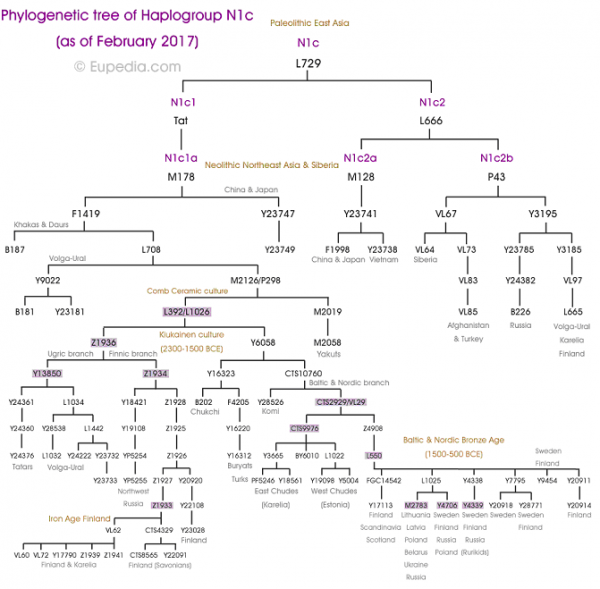 Interestingly, results from uniparentally-inherited markers (mtDNA and Y chromosome) as well as certain phenotypic SNPs also show Siberian signals in Bolshoy: mtDNA haplogroups Z1, C4 and D4, common in modern Siberia24,34,35 are represented by the individuals BOO002, BOO004 and BOO006, respectively (confirming previous findings24), whereas the Y-chromosomal haplotype N1c1a1a (N-L392) is represented by the individuals BOO002 and BOO004. Haplogroup N1c, to which this haplotype belongs, is the major Y-chromosomal lineage in modern north-east Europe and European Russia. It is especially prevalent in Uralic speakers, comprising for example as much as 54% of eastern Finnish male lineages today36. Notably, this is the earliest known occurrence of Y-haplogroup N1c in Fennoscandia. Additionally, within the Bolshoy population, we observe the derived allele of rs3827760 in the EDAR gene, which is found in near-fixation in East Asian and Native American populations today, but is extremely rare elsewhere37, and has been linked to phenotypes related to tooth shape38 and hair morphology39 (Supplementary Data 2). Scandinavian hunter-gatherers from Motala in Sweden have also been found to carry haplotypes associated with this allele4. Finally, in the Bolshoy individuals we also see high frequencies of haplotypes associated with diets rich in poly-unsaturated fatty acids, in the FADS genes4,40,41. |
|
|
|
Post by Admin on Dec 6, 2018 18:00:24 GMT
 The arrival of Siberian ancestry in Europe We formally tested for admixture in north-eastern Europe by calculating f3(Test; Siberian source, European source) statistics. We used several Uralic-speaking populations—Estonians, Saami, Finnish, Mordovians and Hungarians—and Russians as Test populations. Significantly negative f3 values correspond to the Test population being admixed between populations related to the two source populations42. We used multiple European and Siberian sources to capture differences in ancestral composition among proxy populations: As proxies for the Siberian source we used Bolshoy, Mansi and Nganasan, and for the European source we used modern Icelandic, Norwegian, Lithuanian and French. Our results show that all of the test populations are indeed admixed, with the most negative values arising when Nganasan are used as the Siberian source (Supplementary Data 3). Among the European sources, Lithuanians gave the most negative results for Estonians, Russians and Mordovians.  For modern Hungarians, the European source giving most negative results was French, while both Bolshoy and Nganasan gave equally negative results when used as the Siberian source. With Finns as test, modern Icelanders were the European source giving most negative statistics. Finally, Icelanders and Nganasan used as the European and Siberian sources, respectively, yielded the most negative result for the present-day Saami as a Test. This result is still non-significantly negative, either due to the low number of modern Saami individuals in our dataset ( n = 3), or due to post-admixture drift in modern Saami. A high degree of population-specific drift can affect f3-statistics and result in less negative and even positive values42. Indeed, post-admixture drift would correlate well with the suggested founder effect43 in Saami. To further test differential relatedness with Nganasan in European populations and in the ancient individuals in this study, we calculated f4(Mbuti, Nganasan; Lithuanian, Test) (Fig. 3). Consistent with f3-statistics above, all the ancient individuals and modern Finns, Saami, Mordovians and Russians show excess allele sharing with Nganasan when used as Test populations. Of all Uralic-speaking populations in Europe, Hungarians are the only population that shows no evidence of excess allele sharing with Nganasan compared to that of Lithuanians, consistent with their distinct population history from other Uralic populations as evidenced by historical sources (see ref. 44 and references therein). 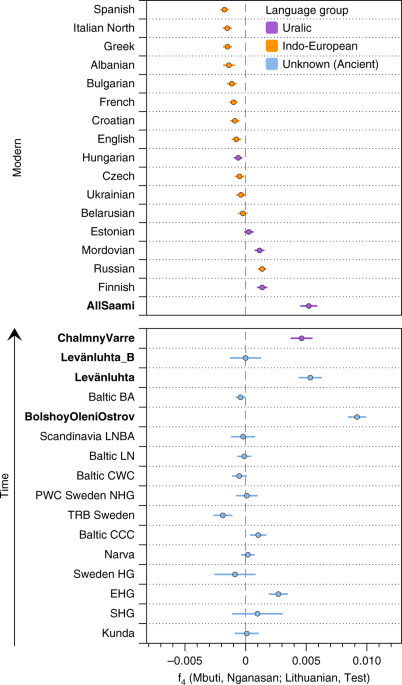 Fig. 3 We further estimated the genetic composition in these populations using qpAdm3. All ancient and modern individuals from the Baltics, Finland and Russia were successfully modelled as a mixture of five lines of ancestry, represented by eastern Mesolithic hunter-gatherers (EHG, from Karelia), Yamnaya from Samara, LBK from the early European Neolithic, western Mesolithic hunter-gatherers (WHG, from Spain, Luxembourg and Hungary), and Nganasan, or subsets of those five (Supplementary Data 4). In contrast to previous models for European populations using three streams of ancestry2,3, we found that some populations modelled here require two additional components: a component related to modern Nganasans, as discussed above, and additional EHG ancestry, not explained by Yamnaya (who have been shown to contain large amounts of EHG ancestry themselves3).  Indeed, the six Bolshoy individuals have substantial amounts of EHG but no Yamnaya ancestry. We find that Nganasan-related ancestry is significantly present in all of our ancient samples except for Levänluhta_B, and in many modern, mainly Uralic-speaking populations. The 3500-year-old ancient individuals from Bolshoy represent the highest proportion of Siberian Nganasan-related ancestry seen in this region so far, and possibly evidence its earliest presence in the western end of the trans-Siberian expanse (Fig. 4). The geographically proximate ancient hunter-gatherers from the Baltics (6000 and 6300 BC) and Motala (~ 6000 BC), who predate Bolshoy, lack this component, as do late Neolithic and Bronze Age individuals from the Baltics7,8,45. All later ancient individuals in this study have lower amounts of Nganasan-related ancestry than Bolshoy (Figs 3, 4), probably as a result of dilution through admixture with other populations from further south. This is also consistent with the increased proportion of early European farmer ancestry related to Neolithic Europeans (Fig. 2b) in our later samples. We note that a low but significant amount of Neolithic European ancestry is also present in the Bolshoy population. Finally, we tested whether Bolshoy, instead of Nganasan, can be used as source population. We found that Bolshoy works as source in some ancient individuals, but not for modern Uralic speakers (see Supplementary Data 4).  Fig. 4 As shown by these multiple lines of evidence, the pattern of genetic ancestry observed in north-eastern Europe is the result of admixture between populations from Siberia and populations from Europe. To obtain a relative date of this admixture, and as an independent line of evidence thereof, we used admixture linkage disequilibrium decay, as implemented in ALDER46. Our ALDER admixture estimate for Bolshoy, using Nganasan and EHG as admixture sources, dates only 17 generations ago. Based on the radiocarbon date for Bolshoy and its uncertainty, and assuming a generation time of 29 years47, we estimate the time of introduction of the Siberian Nganasan-related ancestry in Bolshoy to be 3977 (± 77) years before present (yBP) (Fig. 5b). Estimates obtained using Nganasans and Lithuanians as source populations provided a similar estimate (Supplementary Figure 5 for LD decay plots for multiple populations using Lithuanian and Nganasan as sources.). ALDER provides a relative date estimate for a single-pulse admixture event in generations. When multiple admixture events have occurred, such a single estimate should be interpreted as a (non-arithmetic) average of those events46,48. Therefore the admixture date estimate for Bolshoy does not preclude earlier admixture events bringing Siberian Nganasan-related ancestry into the population, in multiple waves. Indeed, for all other populations with evidence of this ancestry, we find much younger admixture dates (Fig. 5a), suggesting that the observed genetic ancestry in north-eastern Europe is inconsistent with a single-pulse admixture event. |
|
|
|
Post by Admin on Dec 7, 2018 18:02:22 GMT
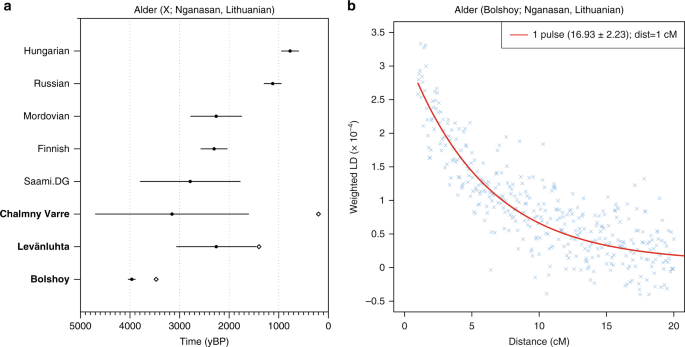 Fig. 5 Major genetic shift in Finland since the Iron Age Besides the early evidence of Siberian ancestry, our ancient samples from Levänluhta and Chalmny Varre allow us to investigate the more recent population history of Finland. To test whether the ancient individuals from Levänluhta form a clade with modern-day Saami or with modern Finns, we calculated f4(Saami(SGDP), Test; X, Mbuti) and f4(Finnish, Test; X, Mbuti), where Test was substituted with each ancient individual from Levänluhta, the two historical Saami individuals from Chalmny Varre, as well as the Modern Saami individual, and X was substituted by worldwide modern-day populations (Supplementary Data 5 & 6, and Supplementary Figures 5 & 6). One Levänluhta (JK1968) and the two Chalmny Varre individuals consistently formed a clade with modern-day Saami, but not with modern-day Finns, with respect to all worldwide populations. One Levänluhta individual (JK1970) showed slightly lower affinity to central Europe than modern-day Saami do, while still rejecting a cladal position with modern-day Finns. This indicates that the people inhabiting Levänluhta during the Iron Age, and possibly other areas in the region as well, were more closely related to modern-day Saami than to present-day Finns; however, their difference from the modern Saami may reflect internal structure within the Saami population or additional admixture into the modern population.  One of the individuals from Levänluhta (JK2065/Levänluhta_B) rejects a cladal position with modern Saami to the exclusion of most modern Eurasian populations. This individual also rejects a cladal position with Finns. We analysed low coverage genomes from four additional individuals of the Levänluhta site using PCA (Supplementary Figure 3), confirming the exclusive position of Levänluhta_B compared to all other six individuals (including the four low-coverage individuals) from that site, as is consistent with the ADMIXTURE and qpAdm results. The outlier position of this individual cannot be explained by modern contamination, since it passed several tests for authentication (see Methods) along with all other ancient individuals. However, no direct dating was available for the Levänluhta material, and we cannot exclude the possibility of a temporal gap between this individual and the other individuals from that site. 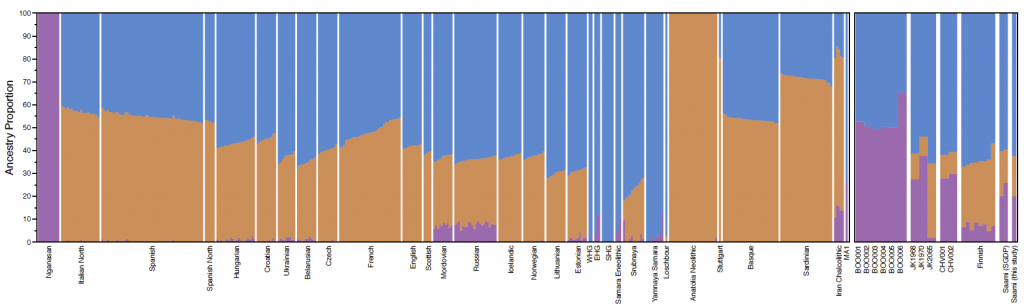 Plot of ADMIXTURE (K=3) results containing West Eurasian populations and the Nganasan. Ancient individuals from this study are represented by thicker bars. The large Nganasan-related component in the Bolshoy individuals from the Kola Peninsula provides the earliest direct genetic evidence for an eastern migration into this region. Such contact is well documented in archaeology, with the introduction of asbestos-mixed Lovozero ceramics during the second millennium BC50, and the spread of even-based arrowheads in Lapland from 1900 BCE51,52. Additionally, the nearest counterparts of Vardøy ceramics, appearing in the area around 1,600-1,300 BCE, can be found on the Taymyr peninsula, much further to the East51,52. Finally, the Imiyakhtakhskaya culture from Yakutia spread to the Kola Peninsula during the same period24,53. Contacts between Siberia and Europe are also recognised in linguistics.  The fact that the Nganasan-related genetic component is consistently shared among Uralic-speaking populations, with the exceptions of absence in Hungarians and presence in the non-Uralic speaking Russians, makes it tempting to equate this genetic component with the spread of Uralic languages in the area. However, such a model may be overly simplistic. First, the presence of the Siberian component on the Kola Peninsula at ca. 3500 yBP predates most linguistic estimates of the spread of extant Uralic languages to the area54. Second, as shown in our analyses, the admixture patterns found in historic and modern Uralic speakers are complex and in fact inconsistent with a single admixture event. Therefore, even if the Siberian genetic component partly spread alongside Uralic languages, some Siberian ancestry may have been already present in the area from earlier admixture events.  Nature Communications volume 9, Article number: 5018 (2018) |
|

