|
|
Post by Admin on Aug 21, 2024 18:52:56 GMT
Fig. 3: Spatial inferences on the origin of Hallstatt individuals. 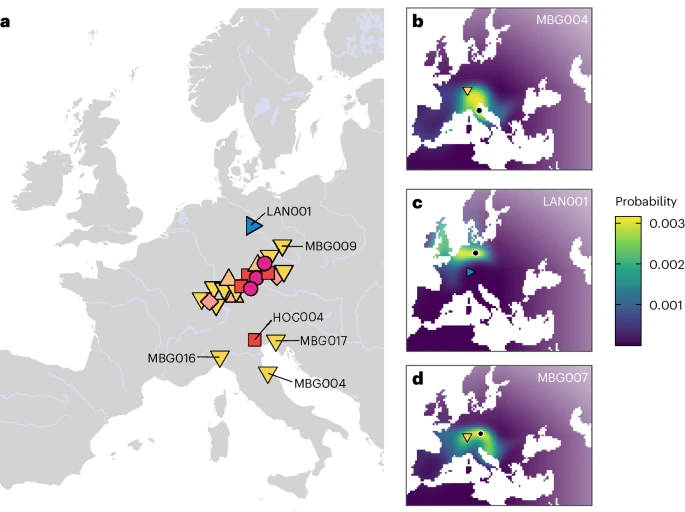 a, MOBEST predictions of the geographic regions where the ancestors of Iron Age individuals (n = 24) from southwestern Germany originated. Shown are the points of maximum probability at search time 0 (the mean date of the respective individual). The symbols and colours correspond to Fig. 1. b, Genetic similarity probability map for MBG004. The filled shape shows the burial location and the black dot the point of maximum probability. c, The same as b for LAN001. d, The same as b for MBG007. Zooming into each site, we reconstruct several biological relationships (third to fourth degree) between the secondary burials MBG001 and MBG013, as well as the three burials MBG002, MBG011 and MBG012 (not indicated in Fig. 1; Supplementary Figs. 2.1 and 2.2), which all show isotopic compositions local to the surroundings of the Magdalenenberg and the Black Forest37 (Supplementary Fig. 2.9). In contrast, none of the secondary burials in Asperg-Grafenbühl and Eberdingen-Hochdorf is related to the respective central graves. Within Asperg-Grafenbühl, we note that the two deceased in the secondary double burial, the adult female APG002 and the male child APG003, are also not biologically related to each other, representing a possible case of fosterage (Discussion). Moreover, APG003 is an outlier in terms of stable isotopes, showing δ18O and 87Sr/86Sr values very similar to the La Tène period male individual LAN001 from a shaft at Alte Burg. While LAN001 most likely originated from coastal northwestern Europe or Central Germany (Supplementary Fig. 2.9), a finding also supported by MOBEST analysis (Fig. 3c), APG003 appears genetically local. His elevated δ18O level may rather reflect breast milk consumption than an origin from a climatically distinct region. Indeed, his 87Sr/86Sr values are very similar to HOC003 and consistent with the biologically available strontium around the Magdalenenberg site, supporting our genetic observation of inter-site mobility. In general, we note that male and female individuals in our sample do not significantly differ in strontium and oxygen isotope values (two-sided Wilcoxon rank-sum exact test; W = 153, P = 0.615 and W = 146, P = 0.4734 for strontium and oxygen, respectively). This stands in contrast to analyses of mobility in Early and Middle Bronze Age southern Germany, where significantly more non-local females than males were found38. Furthermore, we do not find a significant association between grave goods, δ18O and aDNA as markers for non-local origin (Supplementary Note 2). For that, we focused on the Magdalenenberg site where a large number of graves exhibit artefacts of transalpine, south-European (especially North Italian and/or southeast Alpine) provenance37,39, indicating cultural transfer alongside extensive, continuous individual-based mobility. We grouped individuals, for which both isotopes and aDNA data were available, into two groups based on the presence of southern, non-local artefacts. We find that non-local artefacts (being present in 6 out of 16 graves) are not statistically significantly correlated with either higher proportions of EEF ancestry (two-sided Wilcoxon rank-sum exact test; W = 23, P = 0.4923)Fig. 2.11), both indicating cisalpine origin. Consequently, southern grave goods do not constitute a reliable marker of south-European origin in the Magdalenenberg population, although we do identify individuals with such origins in the burial mound via our isotopic and aDNA data. This is especially evident in the case of MBG010, an adult female, who exhibits δ18O and 87Sr/86Sr values indicative of a northern Italian or Iberian origin37 yet is neither buried with southern grave goods nor shows excess genetic affinity to those regions nor δ18O values (two-sided Wilcoxon rank-sum exact test; W = 44.5, P = 0.1283) To supplement our findings on biological relationships, we analysed all individuals for evidence of long runs of homozygosity (RoH), which are indicative of consanguinity (a close biological relationship between the parents). We indeed find two individuals with elevated RoH: MBG004 and APG003 (Supplementary Fig. 2.6). Both individuals exhibit over 150c cM of RoH in total, indicative of recent inbreeding, most likely by first cousin parents40 (Supplementary Fig. 2.7). Given that such high levels are very rare in the published record, the presence of two consanguineous individuals in the comparably small sample size of 30 individuals may suggest that consanguinity was more frequent among the Hallstatt elites of southwestern Germany than in other ancient societies in the archaeogenetic record. |
|
|
|
Post by Admin on Aug 23, 2024 12:20:59 GMT
Emergence and decline of the West-Hallstatt gene pool We compared the genome-wide data of our early Iron Age samples with a reference dataset of 5,665 ancient and 10,176 present-day Eurasian individuals (Methods). When projected on the diversity of present-day Europeans by means of principal component analysis (PCA), we find the Iron Age individuals to be separate in genetic space from present-day Germans and falling closer to present-day French and other southern European individuals (Supplementary Fig. 4.1). Compared with contemporaneous data, the Hallstatt individuals cluster homogeneously intermediate between Iron Age samples from present-day France and the Czech Republic41,42, together with Bronze Age samples from the Bavarian Lech valley38 within the present-day French variation (Supplementary Figs. 4.2 and 4.3). The divergence between prehistoric and present-day individuals from Germany is also seen in the distribution of genetic distances (FST) (Supplementary Fig. 4.4a) as well as correlation of allele frequencies (F4) (Supplementary Tables5.1 and 5.4a). The genetic affinity between our Hallstatt individuals from southern Germany and individuals from Bronze and Iron Age France is part of a broader genetic continuum spanning from Iberia to the Balkan peninsula, featuring a common genetic ancestry component (Fig. 4a, green ‘CWE’ component, Fig. 4b, Supplementary Note 4 and Supplementary Table 4.10). Fig. 4: Population genetic affinities across space and time.  a, Mean supervised ADMIXTURE components at K = 12 (Supplementary Note 4) aggregated across 5,142 individuals from 342 sites dating between 3,150 and 1,750 years BP. b, P values from generalized-likelihood ratio tests implemented in qpWave for testing genetic similarity between southern German Hallstatt individuals and diverse Bronze and Iron Age populations across Europe. Higher P values correspond to higher genetic similarity. c, Overview about population genetic changes in Germany from the Late Neolithic to the present day. The arrows indicate P values from generalized-likelihood ratio tests in qpWave for genetic continuity between temporally preceding and succeeding groups in northern Germany (Lower Saxony, Saxony-Anhalt and Mecklenburg-Vorpommern; symbols in blue) and southern Germany (Baden-Württemberg and Bavaria; symbols in orange), respectively (Supplementary Fig. 5.6). Discontinuities are explicitly marked as interrupted arrows. |
|
|
|
Post by Admin on Aug 25, 2024 18:59:30 GMT
This broad continuum is characterised by a common demographic process, which we see from an analysis of distal ancestry proportions. In particular, using qpAdm we demonstrate an increase of EEF ancestry and a decrease of Yamnaya and Poltavka pastoralists (OldSteppe) ancestry from the Late Neolithic Bell Beaker period onwards, peaking during the Middle Bronze Age and Iron Age (Supplementary Note 4 and Supplementary Figs. 4.4a and 4.5a) and converging the gene pools in France and southern Germany. This increase of EEF is accompanied by a homogenization of the gene pool in terms of EEF and Steppe ancestry, illustrated by a marked decrease of variance of the per-individual statistic F4(YRI, Test; OldSteppe, EEF) between time periods (Supplementary Fig. 4.4d). This phenomenon was described previously38,43 and might reflect continuous admixture with coexisting groups in other regions predominantly from southern Europe, who experienced less gene flow predominantly from southern Europe, who experienced less gene flow from steppe-related populations. It is part of a broader trend of EEF ancestry becoming more similar across central and western Europe in the Bronze Age (Supplementary Fig. 4.5a), coinciding with archaeological evidence of intensified cultural exchange, especially during the Late Bronze Age Urnfield culture period42. Indeed, when estimating the time of admixture in individuals from this region ranging from 2500 to 500 BCE (Supplementary Fig. 4.5d), we observe that admixture time decreases significantly with the date of each individual (Spearman’s rank correlation, P = 2.98 × 10−7), with a slope close to 1.0 (0.75 ± 0.13), which is incompatible with a single pulse of admixture but compatible with stationary continuous and ongoing admixture. To gain insights into the possible sources of this Bronze Age EEF resurgence, we modelled the pooled Hallstatt individuals in qpAdm as a mixture of the Germany_Lech_EBA cluster, and a second source, for which we identify several potential proxies, all of them located in southwestern Europe,especially the Iberian Peninsula and Italy (Supplementary Table 3.12).
To investigate individual ancestries within the Hallstatt group, we used the Middle Bronze Age population from the southern German Lech valley as a proxy for local ancestry. Indeed, most Hallstatt individuals fit a model of receiving all of their ancestry from Germany_Lech_MBA, with the exception of previously described southern outliers MBG004, MBG016 and northern outlier LAN001 from Alte Burg (Supplementary Table 2.8). LAN001 received the majority of his ancestry from a more northern European source, most closely related to the Bronze and Iron Age population of the Netherlands and Saxony-Anhalt (Supplementary Tables 2.9 and 2.11), which is also consistent with his elevated δ18O values supporting a coastal northwestern European or Central German origin44,45,46.
The arrival of individuals of more northern European ancestry during the La Tène period can also be observed in published data from the nearby Czech Republic42, where we analysed individual ancestry components using supervised clustering (Supplementary Fig. 5.8d) and detect a previously undescribed diversification of the gene pool with respect to northern European ancestry from the Hallstatt to the La Tène period (two-sided F test; F = 0.20174, numerator d.f. 15, denominator d.f. 60, P = 0.001). In southern Germany (here Baden-Württemberg and Bavaria) the northern European influx broadens to a major genetic turnover between the Iron Age and the Early Middle Ages (Fig. 4c and Supplementary Note 5). It is illustrated by a sharp decrease of EEF ancestry and a substantial resurgence of Steppe-related ancestry together with a re-diversification of the gene pool (Supplementary Figs. 4.4, 4.5 and 5.2). While the Hallstatt population showed highest genetic affinity to present-day French, Spanish and Belgians, the early medieval (Alemannic and Bavarian) populations of southern Germany47,48 exhibit closest resemblance to present-day Danish, northern Germans, Dutch and Scandinavians (Supplementary Fig. 5.4) and are genetically indistinguishable from Iron Age and Medieval groups in northern Germany and Scandinavia (Supplementary Table 2.10). We argue that this is the result of a major genetic influx from those regions as indicated by qpWave analysis and supervised ADMIXTURE (Fig. 4c and Supplementary Figs. 5.3, 5.5 and 5.6). The northern regions of Germany (here Saxony-Anhalt, Lower Saxony, Mecklenburg-Vorpommern and Schleswig-Holstein) underwent a very different population genetic trajectory than southern Germany. While the Bronze and Iron Age populations in the north also received additional EEF ancestry (Supplementary Figs. 4.5a,b), it was substantially less than what arrived in southern Germany, forming a Steppe ancestry-enriched gene pool highly similar to contemporaneous populations in Denmark, Sweden and Norway (Supplementary Fig. 5.2). Migration from northern Germany introduced EEF-depleted ancestry to southern Germany, resulting in a rise of the median northern European ancestry from 2.8% during the Iron Age to 62.5% during the Early Middle Ages (Supplementary Fig. 5.3), as well as in new paternal ancestry in the form of Y-chromosome haplogroups like I1-M253 (refs. 47,48). While we cannot precisely date this migration, Roman48 and Late Iron Age49 data from Bavaria and Thuringia indicate that parts of the early Iron Age gene pool in southern Germany were not affected until the fourth or fifth century CE (with northern European ancestry not exceeding a median of 8% in these samples). In general, this turnover seems to be part of a larger movement of people, contributing northern European ancestry to the early medieval populations of England50, Hungary51, Italy51 and Spain52.
Most present-day Germans fall between the Hallstatt and early medieval southern German clusters, suggesting a resurgence of EEF-enriched ancestry, especially in southern Germany. This is also indicated by uniparental Y-chromosome evidence. We find that the Hallstatt Y-chromosome gene pool is dominated by R1b-M269 and G2a-P303 lineages, with subhaplogroup G2a-L497 accounting for 37% of the haplotypes in the sample (Supplementary Table 1.1). Interestingly, we find that individuals with haplogroup G2a-L497 (for example, MBG017, MBG016 and HOC004) exhibit significantly more southern European ancestry than individuals carrying haplogroup R1b-M269 (for example, HOC001, APG001 and MBG003) (two-sided Welch two-sample t-test; t = 2.878, d.f. 13.812, P = 0.0123). Although G2a is exceedingly rare in present-day Europe north of the Alps, G2a-L497 still peaks in the area of the former West-Hallstattkreis, namely eastern France, southern Germany, and Switzerland53 as well as northern Italy, thus providing additional evidence for a survival or resurgence of Hallstatt Iron Age ancestry in those regions. Most present-day Germans can be modelled as three-way admixture between SGermany_EIA (54.5 ± 2%), NGermany_Roman (33.8 ± 2.5%) and a third, northeastern European source (here Latvia_BA, 11.7 ± 1.2%) representing further admixture introduced after the initial admixture event, potentially connected to Slavic-speaking populations migrating into eastern Germany during the Middle Ages54 (Supplementary Tables 4.13–14).
|
|
|
|
Post by Admin on Aug 27, 2024 17:45:02 GMT
Discussion
Hereditary leadership is described as one key aspect of early historically recorded complex societies around the world16,55, but it is hard to prove through the archaeological record only. Combining uniparental and autosomal data, we were able to prove a close biological relationship between the two central princely burials of Eberdingen-Hochdorf (HOC001) and Asperg-Grafenbühl (APG001), representing two of the richest graves of European prehistory. Together with dating and osteological estimates of age at death, our pedigree modelling points to a maternal uncle–sororal nephew relationship (most likely model) or a grandfather–daughter–grandson model, suggesting that in this case institutionalized power was matrilineally inherited from the potentate (HOC001), most probably via his sister’s, and less likely via his daughter’s son (APG001). The first and substantially more likely of these scenarios would be congruent with (later) historical Roman accounts of avuncularism among the early Celts of the fifth or fourth century BCE27,56. Today, matrilineally organized societies represent only 12–17% of the world’s populations57, with the majority of societies being patrilineally organized, a pattern also evident from aDNA studies of Neolithic and Bronze Age communities in Europe38,58,59. Yet, global instances of prehistoric societies where hereditary leadership was passed in multigenerational matrilineal descent groups are known60. For Iron Age Europe, matrilineal inheritance of regality is documented for Etruria and Ancient Rome27.
Matrilinear avunculate organization is shown to emerge in populations in which extramarital mating is common and/or paternity confidence is low, so that men are more likely genetically closer related to their sisters’ children than to those of their own wives, ultimately favouring investment in sisters’ children61,62,63,64,65,66. In this context, the observation of inbreeding in two individuals from Asperg-Grafenbühl and Magdalenenberg is indicative. Both individuals are most likely the product of first-cousin mating, a practice often associated with paternity certainty and avunculocal organization, which allows males in matrilineal societies to contribute to sisters’ children who are married to their own wife’s children64,67. In the aDNA record, first cousin mating is exceedingly rare, with less than 3% of ancient individuals showing RoH consistent (but not conclusive) for the offspring of first cousins40. Yet, we highlight that this leadership system may be limited to southern Germany and not apply to the rest of the Hallstatt sphere. In addition, there might be differences between the elite and the larger common population. Recent genetic evidence from the Hallstatt Dolge njive barrow cemetery in Slovenia is neither consistent with a strictly matrilineal nor patrilineal kinship structure for the buried population68 and might indicate a more complex heritability system along both the male and female lines that potentially included adoption or fosterage as well68.
In this context, we find no genetic relationship between the consanguineous Asperg-Grafenbühl child (APG003) and the adult female he was buried with (APG002), nor the main burial, potentially representing an instance of ‘alliance fosterage’69,70,71, a practice associated with the establishment of reciprocal claims on loyalty between status groups and ultimately feudatory state formation72,73. Additionally, a fosterage model would also be supported by his 87Sr/86Sr values, indicating that he originated from the periphery of the Magdalenenberg site, agreeing with written records of non-kinship fostering among the continental and insular Celtic elites70,71.
We find further evidence of familial interconnectedness between the earlier site of Magdalenenberg and the later Eberdingen-Hochdorf in the form of a third-degree genetic relationship between MBG009 and HOC003 and seventh- to eighth-degree relationships between the Magdalenenberg princely burial MBG017, secondary burial MBG016 and HOC004. Together with the relationship between HOC001 and APG001, these connections link the three monumental tumuli. Such instances of non-random mating across a linear geographic distance of more than 100 km and a time span of up to 140 years suggest a high degree of social complexity and the emergence of regional-scale hierarchy. In general, the isotopic profiles of the Magdalenenberg population indicate high, continent-wide mobility during their lifetime and may represent the signature of marital alliance structures and patronal fosterage that connected the distant elite centres and formed the far-reaching social and economic Hallstatt networks37,39.
|
|
|
|
Post by Admin on Aug 29, 2024 19:31:01 GMT
The early Celtic elite of these networks emerged from a long-term population genetic process of ongoing admixture with coexisting groups in southern Europe who previously experienced less gene flow from Steppe-related populations38,42. In this context, we highlight our finding that the earliest elite burial in the region from the central grave of the Magdalenenberg at 616 BCE, as well as his relatives, show evidence of ancestry from South of the Alps, which might suggest a leading role of this connection in the initial formation of the early Celtic Hallstatt culture. Cultural links across the Alps are also preserved in the material culture of these elite graves throughout centuries10,12,39. However, the complex political structures disintegrated in the fifth and fourth century BCE and were ultimately abandoned. Genetic outliers from this and previously published studies suggest that, subsequently, at the height of the Celtic migrations during the fourth and third century BCE, not only ‘Celts’ migrated, but at least a limited number of people from northern central Europe reached the southern zone of the La Tène culture and even northern Italy74, possibly associated with historical entities like the Cimbri and Teutones75. The historical and archaeological record leave no doubt that the development of culture and population in southwestern Germany was temporarily characterized by profound discontinuities, particularly during the third to first century BCE. The definitive end of the 2,000 years of relative genetic continuity from the Bronze throughout the Iron Age in southern Germany is marked by a sudden, sharp increase of Steppe-related ancestry during the Late Antiquity and Early Middle Ages. From a population genetic perspective, this is congruent with arrival of Germanic-speaking tribes from northern Germany or Denmark during the migration period, as also documented by inscription records in the sixth- and seventh-century sites of Baden-Württemberg and Bavaria76. Together with ancestry from eastern Europe introduced during the Middle Ages54, as well as more recent genetic influx from all over the globe, those ancestral populations form the gene pool of the present-day German population.
|
|

