|
|
Post by Admin on Feb 21, 2019 18:30:08 GMT
 A total of 42 reproducible mtDNA sequences (345 bp) were obtained from 62 individual sets of human remains, after discarding 20 samples due to failed amplification or lack of reproducibility. Six of the 42 sequences matched with two archaeologists and one laboratory member were also removed from the study, even though they yielded consistent results through multiple independent extractions. The remaining 36 sequences were inferred to be unambiguous and believable. The following criteria supported the authenticity of the results: (i) an inverse correlation between the size of the PCR amplicons and amplification efficiency (Additional file 4: Table S4); (ii) consistent consensus cloned sequences, although a small number of sites differed from the directly sequenced PCR products, possibly due to random Taq mis-incorporation or DNA damage. Miscoding lesions in clones of PCR products showed that cytosine → thymine changes characteristic of damaged ancient DNA were the most frequent changes in the Xiaohe individuals (Additional file 5: Figure S1); (iii) sex determination by molecular and morphological methods gave consistent results (Table 1); (iv) the mtDNA HVRI sequences corresponded to the key coding region SNPs defined by the mtDNA phylogenetic tree [26]; (v) analysis of cattle bones from the Xiaohe site using the human-specific primers did not reveal human DNA, implying the bones were free of human DNA and the extractions were done cleanly; (vi) the mtDNA sequences from multiple independent DNA extractions and using different samples (tooth, femur) were consistent (Additional file 6: Table S5). The 36 sequences accepted as genuine bone sequences have been submitted to GenBank, with accession numbers KF436896-KF436931. Mitochondrial DNA profiles and haplogroups The 36 successfully typed individuals yielded 21 distinct mtDNA haplotypes, of which 18 could be assigned to 12 previously defined haplogroups [30, 31, 32] by means of HVRI and coding region polymorphisms (Table 1). The haplogroups were the west Eurasian H, K, T, U7, U5a, U2e, the east Eurasian B, C4, C5, D, G2a, and the Indian M5. 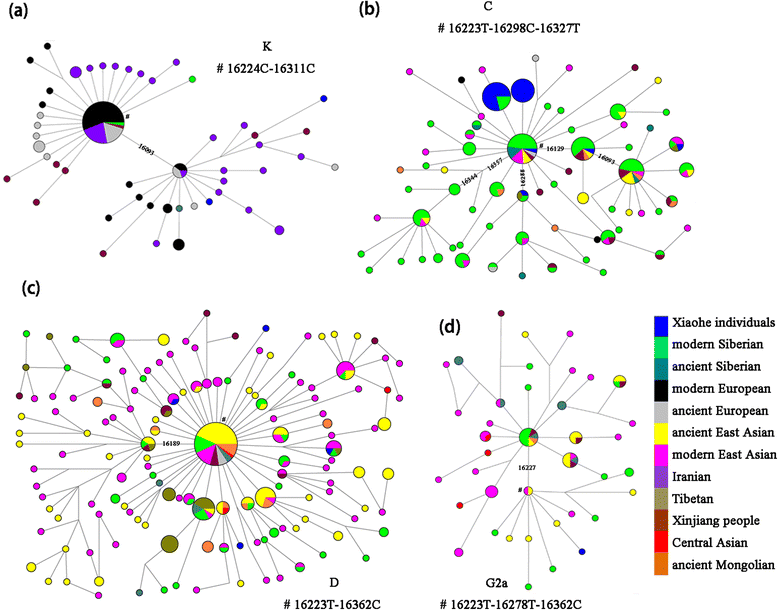 Fig. 3 Median joining networks for mtDNA haplogroups K, C, D and G2a, based on HVS-I sequences between region np16050-16391. Circle areas are proportional to haplotype frequency. The length of the lines between nodes is proportional to the mutation steps. The diagnostic mutations used to classify the major branches are labeled on the line. The Number sign(#) and the following panels indicate the assumed root of each haplogroup The west Eurasian haplogroups of the Xiaohe people were more diverse (Hd = 0.9722 versus Hd = 0.8585), but less abundant (9 individuals versus 26 individuals) than the East Eurasian haplogroups. The predominant lineage was UK, of which four different subhaplogroups were observed: one K, two U7, two U5a, and one U2e. One individual with Hg T and one individual with Hg H were detected. The latter carried the HVRI Cambridge Reference Sequence (CRS), very common in living Europeans [31, 33, 34]. This sequence has also been observed in ancient human remains of Neolithic Europe [35, 36], the Bronze Age in central Asia [37], as well as the Mongolian Altai Mountains [38], and the Iron Age in southern Siberia [39]. The T haplotype observed in Xiaohe is found exclusively in Europeans, with the exception of Iran in modern people, and found mostly as T2. It has also been observed in human remains of Neolithic Europe [36], the Eneolithic/Bronze Age in the Pontic Caspian steppe [40], and the Bronze Age in Kazakhstan [37]. No exact match was found for the Xiaohe K haplotype in our database. The network shows that it clusters into one subclade with the 16093 mutation, which is mainly distributed in Europe and Iran (Fig. 3a). Therefore, the K haplotype sequenced in Xiaohe is currently uninformative about population affinity. There are two U5a haplotypes observed in Xiaohe, the basal U5a*(16192 T-16256 T-16270 T) was found broadly in Europe and central Asia, while the derived U5a haplotype(16192 T-16256 T-16270 T-291 T) was found exclusively in Europe for modern people. These two sequences have also been found in Neolithic Europe [35, 41, 42]. U5a is a very ancient and important European haplogroup and is thought to have expanded eastward into central Siberia. It has been observed in human remains of the Neolithic in the Baikal regions and the Bronze Age in the Altai and Xinjiang [39, 43, 44]. The U2e sequence observed in Xiaohe did not match any sequence in our database, the most matching sequences (showing one to two np differences) were mainly found in Europe. U2e also was an ancient European lineage like U5, and had spread into Central Eurasia in the Bronze Age [31, 39, 44]. The presence of individuals of Hgs H, T, U5a and U2e in Xiaohe indicates maternal lineages with an ultimate origin in Europe. HgU7 is absent in many parts of Europe, but its frequency increases to >4 % in the Near East and up to 5 % in Pakistan, reaching almost 10 % in Iranians, and its highest frequency in Gujarat. U7 haplogroup probably originated in the region between Iran and Indian Gujarat [45, 46, 47]. The U7 variant observed in Xiaohe is currently found mostly in Iran, Europe and the Tibetan plateau. In addition, we found one individual with the Indian lineage M5 [48]. Nowadays, the M5 variant observed in this study is found mainly in south and southwest Asia. The presence of hgs U7 and M5 in the Xiaohe people suggests that populations of west/south Asia contributed to the gene pool of the Tarim Basin during the Bronze Age. The most dominant east Eurasian haplogroup in the Xiaohe people was C, found in 18 of the 36 individuals (47 %) and associated with five distinct mtDNA C4 haplotypes and one C5 haplotype. Nine Xiaohe individuals carried the variant 16223-16298-16309-16327 and five carried the variant 16298–16327. The first of these variants, 16223-16298-16309-16327, has to our knowledge not been previously observed in ancient or living populations, while the variant 16298–16327 was only observed in present-day Siberia, although at low frequencies [49, 50, 51]. A variant characterised by substitutions 16223-16298-16327, observed in one Xiaohe individual, is found widely in present-day Eurasia, with the highest frequency in central/eastern Siberia. It also been detected in a number of ancient individuals, three from Neolithic central Siberia [43], one from northeast Siberia (3600 yBP) [52], six from northeast Europe (3500yBP) [37], twelve from the Bronze Age West Siberian Plain [53], one from southern Xinjing(2800-2011yBP) [54] and four from late Neolithic northwest China [55]. Haplotype 16129-16223-16298-16327 is found mainly in currently northeast, central and south Siberian populations, in Mongolia and central Asia. It also was found in one ancient Mongolian (2000 yBP) [56]. Haplotype 16093-16129-16223-16298-16311-16327 is probably rare, since it has only been detected previously in four present-day individuals, one in south Siberia, one in Tibet, one in Southeast Asia, and one in China. One Xiaohe individual carried Hg C5 (16223-16288-16298-327), of a variant only observed previously in one individual of southern Siberia, and in one of the Tibetan Plateau (Fig. 3b). The second most frequent east Eurasian haplogroup in the Xiaohe people was D, found in four individuals, with four different variants. The first, 16051-16223-16362, is found mainly in Southeast Asia. The second, 16223-16234-16316-16362, is found throughout the Eurasian continent, including China, Japan, Siberia, and Eastern Europe. The remaining two D haplotypes had no exact match in any of the available databases. Interestingly, hg D has been observed at high frequency in Hami people, a Bronze Age population of northeast Xinjiang [44]. It is also been observed in Neolithic Chinese and Siberians [43, 55]. In the Network Tree, We can see that some Xiaohe D haplotypes cluster into the East Asian subclade, the others cluster into the Siberian subclade (Fig. 3c). Therefore, the D haplotype sequenced in Xiaohe is currently uninformative about population affinity. One individual carried G2a, but no matching sequence was found in the database. G2a is relatively abundant in northern China and central Asia, reaching significant levels in Southern Siberia [50]. However, Xiaohe G2a haplotype clusters into one of the East Asian clades in the Network tree (Fig. 3d), indicating close affinities to East Asians. One single individual carried hg B, an important East Asian haplogroup, of a particular variant not previously observed. The presence of haplogroups C4, C5, D, G2a and B in Xiaohe people indicates close affinities to Siberians and East Asians. |
|
|
|
Post by Admin on Feb 21, 2019 20:27:08 GMT
Comparison of the Xiaohe population with ancient and extant populations of Eurasia In order to characterise the genetic relationship between the Xiaohe population and other ancient and extant Eurasian populations, the PCA based on the mtDNA haplogroup frequencies and the MDS plot based on genetic distance between sequences were conducted. However, as many individuals had identical C4 haplotypes, indicating potential maternal relationships within the population, the frequency of C4 was likely to be overestimated. To account for this, we assumed a scenario of extreme maternal kinship, where identical haplotypes in several individuals of the same layer were only counted once. The PCA plot of the first two components showed that present-day populations largely segregate into three main clusters: Europeans, Siberians, and Central/East Asians (Fig. 4). Europeans and Central/East Asians were separated along the first component axis (23.34 % of the variance), reflecting their longitude. Europeans and Siberians were separated along the second component axis (23.04 % of the variance). Xiaohe maternal lineages were closest to the Xinjiang populations (modern Xinjiang population and ancient Hami people), and second-closest to the central Siberians (Tuvinians). An MDS plot confirmed the genetic affinity with Siberians inferred from the PCA, but showed a long distance with Central /East Asians (Additional file 7: Figure S2). 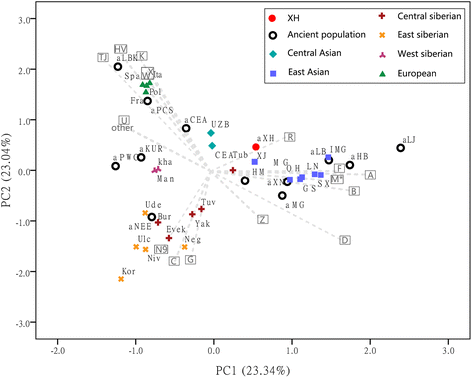 Fig. 4 Principal Component Analysis of mitochondrial haplogroup frequencies. The first two dimensions account for 46.38 % of the total variance. Grey arrows represent haplogroup loading vectors, i.e., the contribution of each haplogroup. Ancient populations included in this study: aXH: Xiaohe cemetery; aCA: Nomads from Kazakhstan (2,100–3,400 yBP); aKur: Siberian Kurgans (1,600–3,800 yBP); aPWC: Scandinavian Pitted-Ware Culture foragers (4,500–5,300 yBP); aLBK: German early Neolithic Linear Pottery Culture population(6,900–7,500 yBP);aNEE: North East European ancient people (3,500–7,500 yBP):aLB: Neolithic Lake Baikal population (6,130–7,140 yBP); aHM: Xinjiang Hami people (4000yBP); aHB: Chinese Shanxi Hengbei people (3000yBP); aMG and aLJ: late Neolithic Qijia Culture peopulions in Ganqing region of China (4000yBP); aXN: nomads from Mongolia (2500yBP). Detailed information on the ancient and modern populations is provided in Additional file 3: Table S Our previous analysis of DNA from the deepest layer of burials of the Xiaohe site revealed that the first settlers had European paternal lineages, and maternal lineages of European and central Siberian origin, consistent with the “steppe hypothesis” of the origins of the first inhabitants of the Tarim Basin [23]. In the present study, analysis of the remaining four, more recent burial layers, confirmed that the origin of the mitochondrial lineages is more widespread, and we detected west Eurasian lineages H, K, U5, U7, U2e, T, east Eurasian lineages B, C4, C5, D, G2a, and Indian lineage M5. Haplotypes H, K, U5 and T are found mostly in Europe, suggesting genetic affinities with Europe. While Xiaohe U2e haplotype has not been observed in living populations, the hg U2e is thought to have originated in Europe, from where it had been spread into central Siberia in the Bronze Age [39]. The distribution of these haplogroups overlaps with the regions of the Afanasievo culture, Andronovo culture or Yamna culture, but is remote from the Oxus civilization. These west Eurasian genetic components in the Xiaohe people corroborate the “steppe hypothesis”. However, layers 1–4 also had individuals with hgs U7 and M5, common in west/south Asian populations today, but rare in Europeans and Siberians. Although the genetic structure of the oasis people in the Bronze Age is unclear, archaeological evidence indicates that settled populations of the oasis civilization in central Asia descended from farmers from the southwest [17]. These ancient central Asians had been in contact with south Asians and likely received a genetic contribution from them. Considering the archaeological materials and the environmental similarities between central Asia and the Tarim Basin, hgs U7 and M5 observed in Xiaohe people more likely originated from the oasis peoples but not directly from west/south Asians. This suggests populations from the oasis may have made a later contribution to the gene pool of the Xiaohe people, giving some credence to the “oasis hypothesis”. The later Xiaohe people (layers 1–4) carried diverse east Asian maternal lineages, including the predominant C4, as well as C5, which has a similar geographical distribution to C4, suggesting links with Siberia, especially central/south Siberian populations. Although hgs B, D and G2a are common in East Asians and Mongolians besides Siberians, except for broomcorn millet (P. miliaceum), there was no archaeological or anthropological evidence in the Xiaohe cemetery for links with East Asia. However, hgs C and D have also been observed in Bronze Age human remains from North Xinjiang (Hami), a place where culture and human features appear to indicate a blend of both east and west. DNA analysis showed that the Hami people had close affinities with Neolithic people in Ganqing region of China [44]. Recently archaeobotanical analysis considered that East Asian domesticated broomcorn likely was introduced into Central Eurasia via the route of North Xinjiang from Ganqing region at middle third millennium BC. Therefore, some eastern components in the later Xiaohe people may have derived from North Xinjiang and have an ultimate origin in East Asia but not central/southern Siberia, something still consistent with the “steppe hypothesis”. This was indicated by the close relationship of the Xiaohe population with populations of Xinjiang in the PCA graph (Fig. 4).  Xiaohe people displays higher and higher levels of haplotype diversity (fifth layer Hd = 0.7381, fourth layer Hd = 0.9004, layers1-3 Hd = 0.9890) from earlier to later, suggesting multiple population incursions into the Tarim Basin after its initial settlement. People carrying European maternal lineages may have spread east into south Siberia, where they mingled with local populations and eventually spread south into Xinjiang via the Ertix River. However, ancient DNA analyses indicate that the west Eurasian lineages observed in ancient south Siberia were associated with the eastward spread of Europeans of the Afanasievo culture [39]. This suggests that the European components could have reached north Xinjiang later, via the Kazakh steppe northwest of the Tarim Basin. Interestingly, the cattle excavated from the Xiaohe cemetery carried mainly lineage T3, typical of European cattle [57]. These diverse lines of evidence support the“steppe hypothesis”. In contrast, people bearing the south /west Asian components could have reached the Tarim Basin through the Pamirs, moving eastward along the south or north edges of the Tarim Basin. Recently one study showed that agricultural populations had contact with nearby mobile pastoralists at the beginning of the second millennium BC in Central Asia [58], indicating that genetic components of agriculturalists might also introgress into pastoralist populations. This was confirmed by the evidence that one Indian haplogroup was found in ancient Kazakhstan [37]. Therefore, people bearing the south/west Asian components could have first married into pastoralist populations, and reached North Xinjiang through the Kazakh steppe following the movement of pastoralist populations, then spread from north Xinjiang southward into the Tarim Basin across the Tianshan Mountains, and intermarried with the earlier inhabitants of the region, giving rise to the later, admixed Xiaohe community. Given that the south/west Asian components are relatively minor in the Xiaohe population, it is likely that nomadic herders from northern steppe had a greater impact on the eastern Tarim Basin than the Central Asian oasis farmers. The archaeological evidence for woolen textiles and the medicinal plant Ephedra in the earliest Xiaohe layer and the Gumugou site indicate that the oasis culture had reached the Tarim Basin in the early Bronze Age. It is well known that Ephedra was used by oasis farmers, whereas it does not grow in the Russo-Kazakh steppe, nor is associated with the Afanasievo or Andronovo cultures [5, 7]. Furthermore, the wheat excavated from Xiaohe was hexaploid bread wheat, a cereal grain cultivated originally in the Near East [59]. Therefore, it is possible that the oasis route may have been significant in the peopling of Xinjiang in the early Bronze Age, at least northern or western Xinjiang. This was supported by the evidence that Indian haplogroup M25 was observed in one ancient individual from later Neolithic Ganqing region (data unpublished). The groups reaching the Tarim Basin through the oasis route may have interacted culturally with earlier populations from the steppe, with limited gene flow, resulting in a small genetic signal of the oasis agriculturalists in the Xiaohe community. BMC Genetics 2015 16:78 |
|
|
|
Post by Admin on Feb 23, 2019 17:35:28 GMT
 The Tarim Basin in western China, positioned at a critical site on the ancient Silk Road, has played a significant role in the history of human migration, cultural developments and communications between the East and the West. It became famous due to the discovery of many well-preserved mummies within the area. These mummies, especially the prehistoric Bronze Age 'Caucasoid' mummies, such as the 'Beauty of Loulan', have attracted extensive interest among scientists regarding who were these people and where did they come from. Based on analyses of human remains and other archaeological materials from the ancient cemeteries (dated from approximately the Bronze Age to the Iron Age), there is now widespread acceptance that the first residents of the Tarim Basin came from the West. This was followed, in stages, by the arrival of Eastern people following the Han Dynasty [1, 2]. However, the exact time when the admixture of the East and the West occurred in this area is still obscure [3]. In 2000, the Xinjiang Archaeological Institute rediscovered a very important Bronze Age site, the Xiaohe cemetery, by utilizing a device employing the global positioning system. The rediscovery of this cemetery provided an invaluable opportunity to further investigate the migrations of ancient populations in the region. The Xiaohe cemetery (40°20'11"N, 88°40'20.3"E) is located in the Taklamakan Desert of northwest China, about 60 km south of the Peacock River and 175 km west of the ancient city of Kroraina (now Loulan; Figure 1). It was first explored in 1934 by Folke Bergman, a Swedish archaeologist, but the cemetery was lost sight of until the Xinjiang Archaeological Institute rediscovered it in 2000. The burial site comprises a total of 167 graves. Many enigmatic features of these graves, such as the pervasive use of sexual symbolism represented by tremendous numbers of huge phallus-posts and vulvae-posts, exaggerated wooden sculptures of human figures and masks, well-preserved boat coffins and mummies, a large number of textiles, ornaments and other artifacts, show that the civilization revealed at Xiaohe is different from any other archaeological site of the same period anywhere in the world [3]. 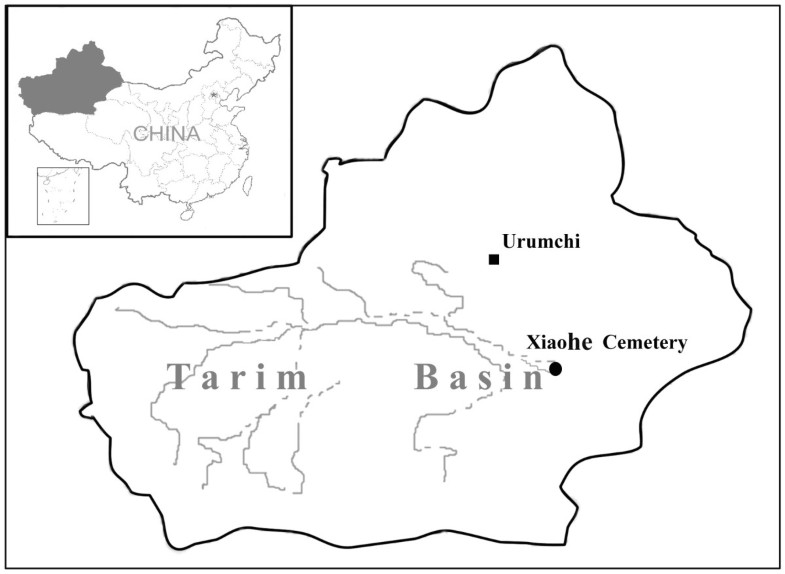 The entire necropolis can be divided, based on the archeological materials, into earlier and later layers. Radiocarbon measurement (14C) dates the lowest layer of occupation to around 3980 ± 40 BP (personal communications; calibrated and measured by Wu Xiaohong, Head of the Laboratory of Accelerator Mass Spectrometry, Peking University), which is older than that of the Gumugou cemetery (dated to 3800). To date, these are the oldest human remains that have been excavated in the Tarim Basin [3]. A genetic study of these invaluable archeological materials will undoubtedly provide significant insights into the origins of the people of the Tarim Basin. We examined the DNA profiles on both the maternal and the paternal aspects for all the morphologically well-preserved human remains from the lowest layer of the Xiaohe cemetery. We used these data to determine the population origins, to provide insights into the early human migration events in the Tarim Basin and, finally, to offer an expanded understanding of the human history of Eurasia. |
|
|
|
Post by Admin on Feb 24, 2019 17:30:35 GMT
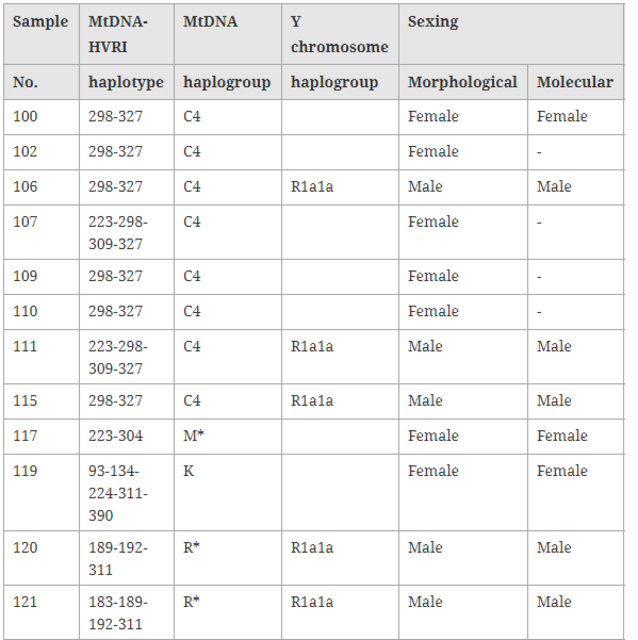
MtDNA haplogroup profile and distribution The 20 mtDNA fragments containing eight different haplotypes that can be further assigned to five haplogroups, which belong to subhaplogroups of macrohaplogroups M and N (Figure 2). 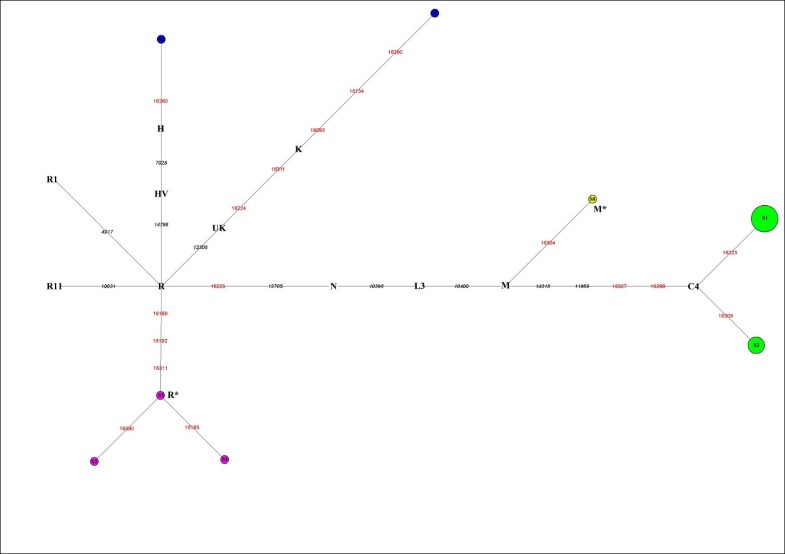 Figure 2 Reduced median network of Xiaohe sequences. Node size is proportional to frequencies. HVR1 positions are numbered relative to Cambridge reference sequence (CRS). Single nucleotide polymorphism diagnostic positions are in black italics; green represents East Eurasian lineage C, containing 14 individuals. Xiaohe R* is the cluster under the macrohaplogroup R. The dominant haplogroup in the Xiaohe people was the East Eurasian lineage C, shared by 14 Xiaohe individuals who were associated with two different mtDNA haplotypes (S1 and S2). According to the coding region 11969 G to A, all lineage C found in the Xiaohe people was further changed to subhaplogroup C4, which had D-loop group-specific polymorphisms at nucleotide positions (np) 16298 (T to C) and 16327 (C to T) [18]. Interestingly, the haplotype S1 shared by these 10 of the 14 Xiaohe individuals did not have the 16223T, a mutation had been found in the majority of modern lineage C populations. This haplotype was found merely in modern Evenks and Udegeys of southeastern Siberia but the frequency is low [18]. It is interesting to note that our study found a mutation, unique to the Xiaohe people but rare among lineage C, 16309 G (see Additional File 3). To the best of our knowledge, the lineage C with 16309 G was observed only in three people from modern central Asia to date, who also possess another two mutations in HVRI region [19].  Besides the East Eurasian lineage, two West Eurasian mtDNA haplogroups H and K were found among the Xiaohe people. H lineage is the most common mtDNA haplogroup in West Eurasia [20], but haplogroup H with a 16260T was shared by only nine modern people in Genbank, including one Italian, one German, one Hungarian, one Portuguese, one Icelander and four English people. Haplogroup K, a western Eurasian-specific haplogroup, is mainly distributed in Europe, central Asia, and Iran [20, 21]. However, haplogroup K with 16134T, found in the Xiaohe people, has not been found in modern people to our knowledge. Among the Xiaohe people, three sequences with the unique HVRI motif 16189-16192-16311 formed a subcluster (Figure 2) and were not shared by modern people. They are identified as macrohaplogroup R through sequencing the PCR amplicons at np10400 and np12705 in the coding region. The np12308, np14766, np10031, np4917, np3970, and 9 bp deletion, which are the diagnostic sites for the main subhaplogroups of R, were further examined [15]. The results showed that they are related neither to the West Eurasian haplogroups UK, TJ, HV, R11 and R1, nor to the East Eurasian haplogroups B and F. So we designated them as haplogroup R* temporarily. Another sequence with motif 16223-16304, shared by some people from East Asia, India, and Europe, was assigned to haplogroup M*. 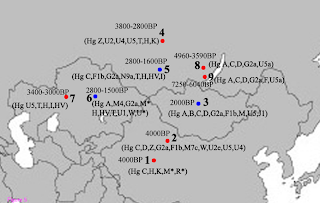 Y chromosome haplogroup profiling and distribution Fifteen individuals' AMG amplicons were obtained from the 20 Xiaohe individuals (whose mtDNA was successfully amplified), among which seven individuals were identified as male and eight as female. The Y chromosome haplogroup of the seven males were all assigned to haplogroup R1a1a through screening the Y-SNPs at M89, M9, M45, M173 and M198 successively. Haplogroup R1a1a is widely distributed in Eurasia: it is mainly found in Eastern Europe, Central Asia, South Asia, Siberia, ancient Siberia, but rare in East Asia [22, 23, 24]. |
|
|
|
Post by Admin on Feb 25, 2019 18:24:28 GMT
The Xiaohe cemetery is the oldest archeological site with human remains discovered in the Tarim Basin to date. Our genetic analyses revealed that the maternal lineages of the Xiaohe people were originated from both the East and the West, whereas paternal lineages discovered in the Xiaohe people all originated from the West. The East Eurasian lineage C, which was widely distributed in modern Asian populations, was the dominant haplogroup in the remains recovered from the lowest layer of the Xiaohe cemetery. This lineage is most frequently found in modern Siberian populations (Evenks, Yakut, Evens, Tuvinian, Buryat, Koryak and Chukchi) and to a lesser extent in modern East Asian (Mongolian, Daur and Korean) and Central Asian populations [25, 26, 27, 28, 29] (Figure 3). It was also found in the ancient Qinghai (4000BP) of China [30] and ancient South Siberian populations [31, 32]. In order to trace the original wellspring of lineage C in the Xiaohe population, a phylogenetic tree was constructed using 14 ancient Xiaohe samples and 522 modern haplogroup C samples from surrounding areas of the Xiaohe cemetery, including Siberian, Mongolian, Central Asian, northern Chinese and northern minorities of East Asia (see Additional File 3). The phylogenetic network displays a star-like distribution within the South Siberian population, which has an ancestral haplotype motif 16223-16298-16327. The ancestral haplotype was found mainly in South Siberian whose diversity of haplotypes C is very high (Figure 4). Therefore, the original source of haplogroup C was inferred to South Siberian. It is important to note that the C haplotypes of the Xiaohe people had only a single mutation compared with the ancestral haplotype. The shared sequences of the Xiaohe C haplotype (S1) were distributed in southeastern Siberia. It implies that the east Eurasian component in the Xiaohe people originated from the Siberian populations, especially the southern or eastern Siberian populations. 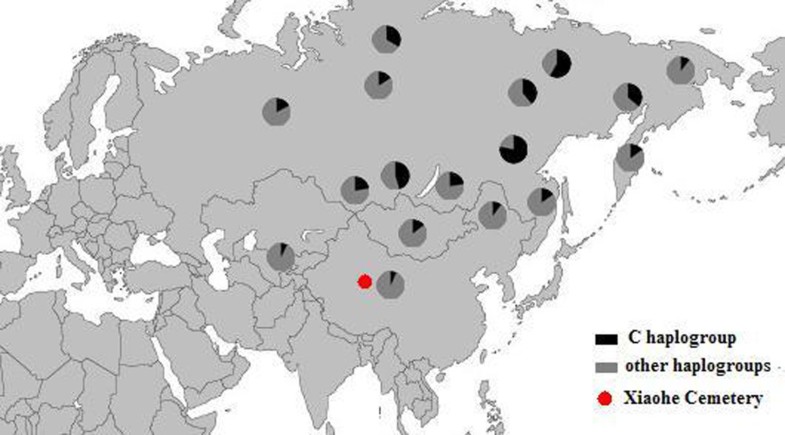 Figure 3 Map of Eurasia, showing the approximate distribution of haplogroup C. Black sections reflect the frequency of haplogroup C data taken from references listed in Additional File 3. The mtDNA haplogroup H is the most common mtDNA haplogroup in Europe, especially in northwestern Europe, and its frequency can be as high as 65% in Iberia. Frequencies gradually decrease from the northwest to the southeast of Europe. By contrast, the frequency of haplogroup H rises to only 20% in the Near East, and to less than10% in Central Asia, and is very low in East Asia [33, 34]. All of the shared sequences of the Xiaohe H haplotype, however, were distributed in Western Europe. Haplogroup K is also common in Europe, particularly around the Alps and the British Isles. It is found with less frequency in North Africa, the Middle East, and South Asia [21, 35, 36, 37]. Considering the presence of haplogroups H and K in the Xiaohe people and the geographical distribution of shared sequences, we conclude that the west Eurasian component observed in the Xiaohe people originated from western European, and maternal ancestry of the Xiaohe people might have close relationship with western European. Regarding the Y chromosomal DNA analyses, the seven males identified all belonged to haplogroup R1a1a. It is most frequently found in Eastern Europe, South Asia and Siberia. In contrast, it is relatively uncommon in Middle Easterners and rare in East Asian [22, 23, 24]. It is thought to be a trace of the migration events of early Indo-European [38, 39]. The presence of haplogroup R1a1a in the ancient Xiaohe people implies that the parental ancestry of the Xiaohe people originated from somewhere in Siberia or Europe, which is consistent with the origin of maternal ancestry. 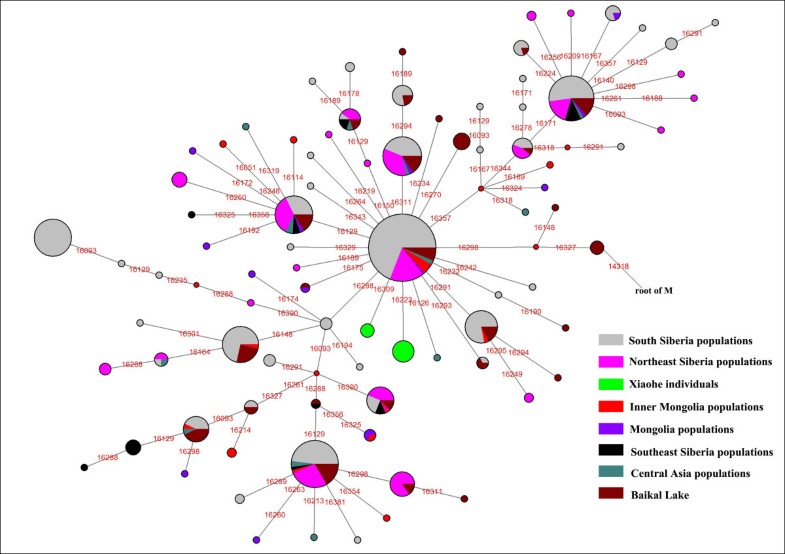 Figure 4 Phylogenetic tree of haplogroup C, based on HVS-I sequences in the region 16050-16391. For references for the mitochondrial DNA sequences in this study see Additional File 3; the length difference mutations were excluded from this analysis. It is generally agreed that the origin of modern populations in Xinjiang and Central Asia is the result of the admixture of people from the West and the East [19, 25, 40]. When and where this admixture first occurred has long been of interest to geneticists and archaeologists [41, 42, 43, 44]. The year 132 BC is often considered to be the beginning of contact between the East and the West along the Great Silk Road, since the Chinese explorer Zhang Qian went westward into Central Asia at that time. However, Mair has suggested that the date should be even earlier, based on the fact that silk appeared in Europe at 1000 BC [1]. In this study, the East and West Eurasian lineages are seen to coexist in the Xiaohe people, implying that the East had contacted the West during the early Bronze Age. It is noteworthy that the maternal lineage of five male individuals (106, 111, 115, 136 and 139) originated from East Eurasian, whereas their paternal lineage originated from the West Eurasian, implying that the Xiaohe population had been an admixture of people from both the West and the East. Given the unique genetic haplotypes and the particular archaeological culture, the time of this admixture could be much earlier than the time at which the Xiaohe people were living at the site. This means that the time of their mingling was at least a 1000 years earlier than previously proposed. However, the mtDNA haplogroups H, K and C all are very ancient lineages, over 10,000 years old in vast north Eurasia, whereas the civilization of the Tarim Basin, according to the archaeological materials, arose very late. The admixture therefore probably occurred elsewhere, before immigration into the Tarim Basin. The Xiaohe people might well have been an admixture at the time of their arrival. Where did the initial admixture occur? 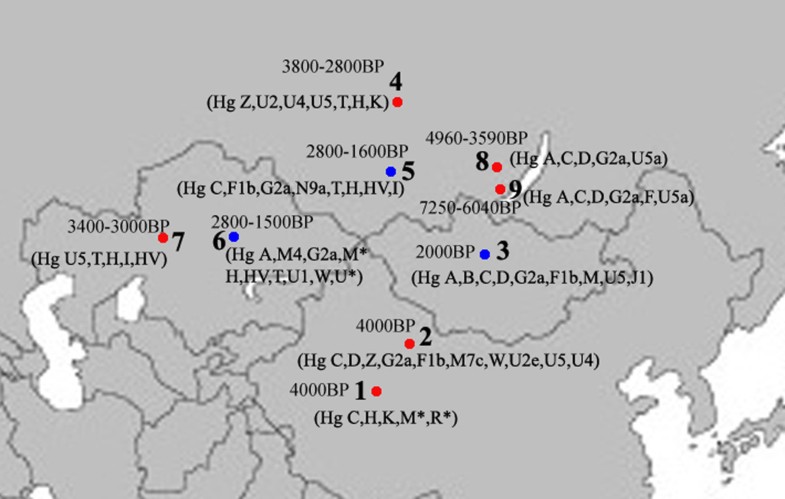 Figure 5 Map of Eurasia, showing ancient populations from the Tarim Basin and surroundings. Number 1 represents Xiaohe cemetery, data from this study; number 2 represents Xinjing Hami cemetery, data not published; Number 3 represents ancient Xiongnu, data from reference 46; Numbers 4 and 5 represent ancient South Siberian people, data from reference 38, Numbers 6 and 7 represent ancient Central Asians, data from reference 41; Numbers 8 and 9 represent ancient Lake Baikal people, data from reference 45. The red colour represents that the data was generated from samples from about Bronze Age and/or the prehistory era, while blue represents that the data was generated from samples from Iron Age. The admixture from people of the West and the East was also found in ancient Central Asia, Siberia, and Mongolia [16, 38, 45, 46]. The extent of the admixture varied in different regions and at different periods (Figure 5). Central Asia has always been the crossroads of contact between the West and the East. Lalueza-Fox et al. proved that Eastern lineages coexisted with Western lineages in Central Asia after 700 BC [41], whereas the West had met the East in south Siberia in the Bronze Age [38], and even earlier at Lake Baikal [45]. Xinjiang and the surrounding areas, especially south Siberia, were places at which the contact between western and eastern populations occurred earlier than in Central Asia. Given the fact that the mtDNA haplogroup C was distributed mainly in south Siberia, and that haplogroups H, K and R1a1a already had spread eastward into south Siberia during the Bronze Age, it is possible that the initial admixture occurred somewhere in southern Siberia. Considering that the cultural characteristics of the Xiaohe cemetery are similar to those of the Andronovo or Afanasevo culture that appeared throughout the southern Russian steppe, Kazakhstan, and western Central Asia during the second millennium BC [1, 46], the admixed population might have had relationship with populations settled South Siberia during the Bronze Age. BMC Biology20108:15 |
|

