|
|
Post by Admin on Mar 14, 2019 20:38:36 GMT
 Fig. 1. Overview of the ancient Iberian genetic time transect. Genomics of the Iberian Peninsula Ancient DNA studies have begun to help us understand the genetic history and movements of people across the globe. Focusing on the Iberian Peninsula, Olalde et al. report genome-wide data from 271 ancient individuals from Iberia (see the Perspective by Vander Linden). The findings provide a comprehensive genetic time transect of the region. Linguistics analysis and genetic analysis of archaeological human remains dating from about 7000 years ago to the present elucidate the genetic impact of prehistoric and historic migrations from Europe and North Africa. Science, this issue p. 1230; see also p. 1153 Abstract We assembled genome-wide data from 271 ancient Iberians, of whom 176 are from the largely unsampled period after 2000 BCE, thereby providing a high-resolution time transect of the Iberian Peninsula. We document high genetic substructure between northwestern and southeastern hunter-gatherers before the spread of farming. We reveal sporadic contacts between Iberia and North Africa by ~2500 BCE and, by ~2000 BCE, the replacement of 40% of Iberia’s ancestry and nearly 100% of its Y-chromosomes by people with Steppe ancestry. We show that, in the Iron Age, Steppe ancestry had spread not only into Indo-European–speaking regions but also into non-Indo-European–speaking ones, and we reveal that present-day Basques are best described as a typical Iron Age population without the admixture events that later affected the rest of Iberia. Additionally, we document how, beginning at least in the Roman period, the ancestry of the peninsula was transformed by gene flow from North Africa and the eastern Mediterranean. 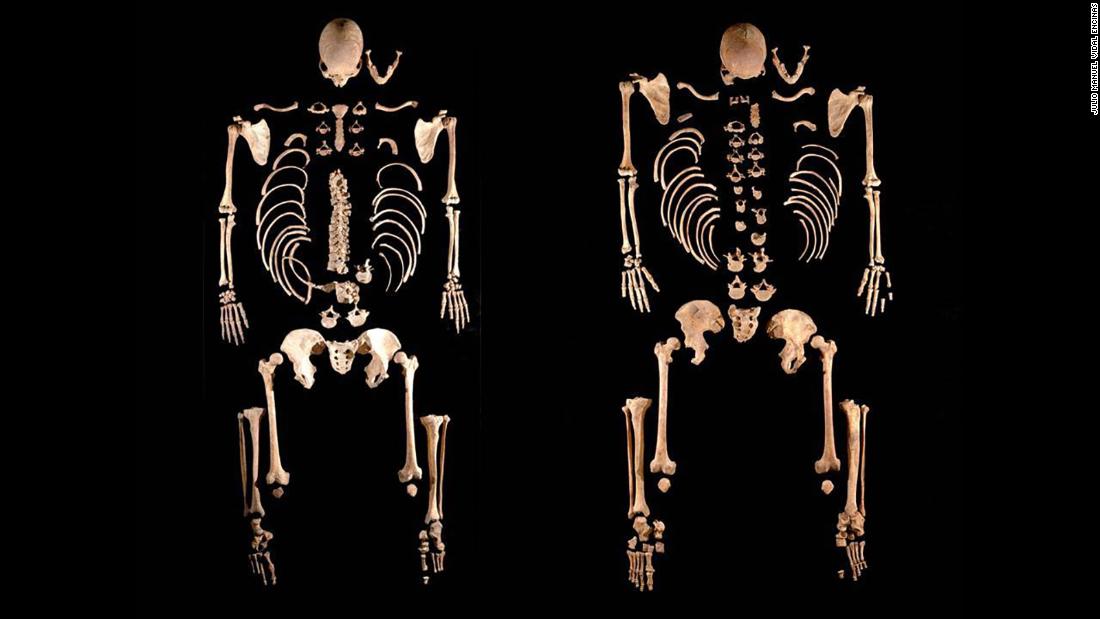 A study published Thursday in the journal Current Biology analyzed the Iberian hunter-gatherer population between 6,000 and 13,000 years ago, and one published in the journal Science conducted an overview of the Iberian population over the past 8,000 years. The findings support some ideas about the area's history while challenging others. The researchers were surprised to discover that local men living on the Iberian Peninsula during the Bronze Age were replaced in the gene pool, their Y chromosomes supplanted by those of men who migrated to the area. "This is one of the strongest pieces of evidence in ancient-DNA research of sex bias in the prehistoric period," said Iñigo Olalde, study author and postdoctoral fellow at Harvard Medical School, in a statement.  Fig. 2. Genome-wide admixture proportions using qpAdm. Beginning in 2500 BC, the researchers discovered, Iberians were joined by people from central Europe whose showed genetic ancestry from the Russian steppe. And over a few hundred years, the locals and the central Europeans interbred. Before this, there is no evidence that locals came into contact with anyone from outside the area. But that changed after 2000 BC, when 40% of Iberian ancestry and 100% of the fathers in the study could be traced back to central Europe. The difference was obvious in the DNA. The remains of a man and a woman buried together at a Spanish Bronze Age site called Castillejo de Bonete showed that the woman was a local and the man's most recent ancestors had come from central Europe. "The results were astonishing," Carles Lalueza-Fox, co-senior study author and principal investigator of the Paleogenomics Lab at the Institute of Evolutionary Biology, said in a statement. "The data suggest there was a major genetic change that is not obvious from the archaeological record."  However, Olalde noted, "It would be a mistake to jump to the conclusion that Iberian men were killed or forcibly displaced as the archaeological record gives no clear evidence of a burst of violence in this period." The genomes of 403 ancient Iberians who lived between 6000 BC and AD 1600, 975 ancient people from other areas and 2,900 current people were analyzed. Modern-day Iberian men can still trace their paternal ancestry to these central Europeans. But the genetic data will need a boost from what anthropology and archaeology can show about the underlying causes for why this Y chromosome shift happened, the researchers said. The researchers also discovered that between 8000 BC and 5500 BC, Iberia's hunter-gatherers were genetically different from each other. This suggests that they interacted with a different group of hunter-gatherers during the Mesolithic era before Anatolian farmers, or those from what is presently Turkey, moved to Iberia and transformed the area yet again. The farmers also mixed with hunter-gatherers, according to their DNA. They uncovered another surprise. The DNA of one person they studied, buried in Iberia between 2400 and 2000 BC, had North African ancestry. Another who lived between 2000 and 1600 BC showed North African ancestry from a grandparent.  At the time, trade existed between Iberia and North Africa, but now the researchers know that people moved between Africa and Europe as well. The gene flow from North Africa and the eastern Mediterranean is apparent in the DNA when Greek and Roman settlements appeared in Iberia. Those multiethnic settlements transformed Iberia and its population even more. The research also sheds light on why the language and culture of present-day Basques are so distinct from those of Iberians. The modern people of Basque Country, in northern Spain, are genetically similar to the Iberian Iron Age people with ancestry from the Russian steppe. While people around them mixed with different groups and changed, the Basques held on to their heritage. In the southeast, we recovered genomic data from 45 individuals dated between the 3rd and 16th centuries CE. All analyzed individuals fell outside the genetic variation of preceding Iberian Iron Age populations (Fig. 1, C and D, and fig. S3) and harbored ancestry from both Southern European and North African populations (Fig. 2D), as well as additional Levantine-related ancestry that could potentially reflect ancestry from Jewish groups (21). These results demonstrate that by the Roman period, southern Iberia had experienced a major influx of North African ancestry, probably related to the well-known mobility patterns during the Roman Empire (22) or to the earlier Phoenician-Punic presence (23); the latter is also supported by the observation of the Phoenician-associated Ychromosome J2. 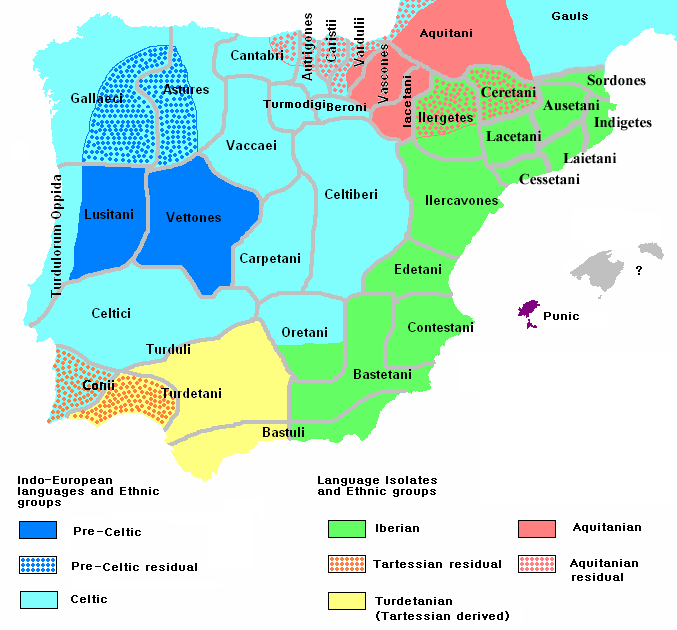 North African ancestry was more widespread in Iberia during the Roman period (about 20 B.C. to 400 A.D.) than previously appreciated, especially in the south. The genetic influences occurred well before groups from North Africa conquered Iberia during the 8th century A.D. Gene flow from North Africa continued into the Muslim period, as is clear from Muslim burials with elevated North African and sub-Saharan African ancestry (Fig. 2D, fig. S4, and table S22) and from uniparental markers typical of North Africa not present among pre-Islamic individuals (Fig. 2D and fig. S11). Present-day populations from southern Iberia harbor less North African ancestry (25) than the ancient Muslim burials, plausibly reflecting expulsion of moriscos (former Muslims converted to Christianity) and repopulation from the north, as supported by historical sources and genetic analysis of present-day groups (25). The impact of Muslim rule is also evident in northeast Iberia in seven individuals from Sant Julià de Ramis from the 8th to 12th centuries CE who, unlike previous ancient individuals from the same region, show North African– related ancestry (Fig. 2C and table S19) and a complete overlap in PCA with present-day Iberians (Fig. 1D). |
|
|
|
Post by Admin on Mar 14, 2019 21:05:46 GMT
 The largest study to date of ancient DNA from the Iberian Peninsula (modern-day Portugal and Spain) offers new insights into the populations that lived in this region over the last 8,000 years. The most startling discovery suggests that local Y chromosomes were almost completely replaced during the Bronze Age. “It's amazing how the technology of ancient DNA, when combined with information from archaeology, anthropology, linguistics and the study of historical records, can bring the past to life,” said Reich, professor of genetics in the HMS Blavatnik Institute, an investigator of the Howard Hughes Medical Institute and co-senior author of the study. The team analyzed genomes from 403 ancient Iberians who lived between about 6000 B.C. and 1600 A.D., 975 ancient people from outside Iberia and about 2,900 present-day people. 271 of the ancient Iberian genomes had not been published before. Nearly two-thirds came from skeletons no older than 2000 B.C., boosting by 25 times the number of publicly available genomes from this relatively recent period. 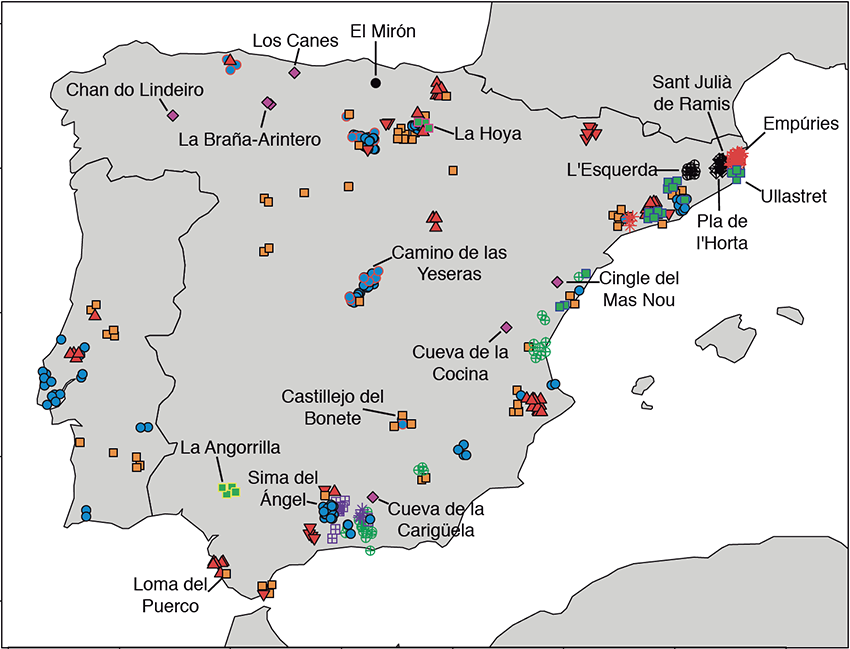 As far back as 2500 B.C., the researchers found, Iberians began living alongside people who moved in from central Europe and carried recent genetic ancestry from the Russian steppe. Within a few hundred years, analyses showed, the two groups had extensively interbred. For example, at a Bronze Age site known as the Castillejo de Bonete in Spain where a woman and man were found buried side by side, analyses revealed that the woman's ancestry was entirely local, while the man had very recent ancestors from central Europe. From the Bronze Age (~2200–900 BCE), we increase the available dataset (6, 7, 17) from 7 to 60 individuals and show how ancestry from the Pontic-Caspian steppe (Steppe ancestry) appeared throughout Iberia in this period (Fig. 1, C and D), albeit with less impact in the south (table S13). The earliest evidence is in 14 individuals dated to ~2500–2000 BCE who coexisted with local people without Steppe ancestry (Fig. 2B). These groups lived in close proximity and admixed to form the Bronze Age population after 2000 BCE with ~40% ancestry from incoming groups (Fig. 2B and fig. S6). Y-chromosome turnover was even more pronounced (Fig. 2B), as the lineages common in Copper Age Iberia (I2, G2, and H) were almost completely replaced by one lineage, R1bM269. These patterns point to a higher contribution of incoming males than females, also supported by a lower proportion of nonlocal ancestry on the X-chromosome (table S14 and fig. S7), a paradigm that can be exemplified by a Bronze Age tomb from Castillejo del Bonete containing a male with Steppe ancestry and a female with ancestry similar to Copper Age Iberians. Although ancient DNA can document that sex-biased admixture occurred, archaeological and anthropological research will be needed to understand the processes that generated it. For the Iron Age, we document a consistent trend of increased ancestry related to Northern and Central European populations with respect to the preceding Bronze Age (Figs. 1, C and D, and 2B). The increase was 10 to 19% (95% confidence intervals given here and in the percentages that follow) in 15 individuals along the Mediterranean coast where non-Indo-European Iberian languages were spoken; 11 to 31% in two individuals at the Tartessian site of La Angorrilla in the southwest with uncertain language attribution; and 28 to 43% in three individuals at La Hoya in the north where Indo-European Celtiberian languages were likely spoken (fig. S6 and tables S11 and S12). This trend documents gene flow into Iberia during the Late Bronze Age or Early Iron Age, possibly associated with the introduction of the Urnfield tradition (18). To the researchers’ surprise, men and women from the two groups contributed strikingly unequal proportions of DNA to subsequent generations. Before the central Europeans moved in, Iberians had no detectable recent ancestry from outside the Iberian Peninsula. After 2000 B.C., 40 percent of Iberians' overall ancestors and 100 percent of their patrilineal ancestors—that is, their father and their father’s father and so forth—could be traced to the incoming groups from central Europe.  Other findings included: Groups of hunter-gatherers scattered across Iberia had markedly different genetic makeup from one another in the Mesolithic era, from about 8000 B.C. to 5500 B.C., suggesting that new groups of hunter-gatherers migrated to Iberia and transformed local populations before farmers arrived with their own distinct ancestry from Anatolia (present-day Turkey). An independent study of Iberian hunter-gatherers, published in Current Biology on the same day as the Science paper, reaches similar conclusions. One person buried in Iberia between 2400 B.C. and 2000 B.C. had entirely North African ancestry, and a second person who lived between 2000 B.C. and 1600 B.C. had a grandparent with North African ancestry. Researchers have known that the two regions engaged in trade at that time; the new study confirms that people also moved from Africa to Europe.  Samples from Iron Age people who lived around 900 B.C. to 19 B.C. in areas where very different languages were spoken all had significant ancestry from the Russian steppe. This suggests that influxes of people did not always spark linguistic changes, specifically the adoption of Indo-European languages, the study authors said. Modern-day Basques are similar genetically to Iron Age people throughout the Iberian Peninsula, leading the researchers to hypothesize that Basque ancestry and language remained relatively intact in the intervening millennia while other groups around them mixed and more significantly changed. In contrast to the demographic changes in the Classical period, movements into Iberia during the decline of the Roman Empire had less long-term demographic impact. Nevertheless, individual sites—for example, the 6th century site of Pla de l'Horta in the northeast—bear witness to events in this period. These individuals, archaeologically interpreted as Visigoths, are shifted from those at L'Esquerda in the direction of Northern and Central Europe (Figs. 1D and 2C and table S18), and we observe the Asian mitochondrial DNA (mtDNA) haplogroup C4a1a also found in Early Medieval Bavaria (20), supporting a recent link to groups with ancestry originally derived from Central and Eastern Europe.
Supplementary Materials for
The genomic history of the Iberian Peninsula over the past 8000 years |
|
|
|
Post by Admin on Mar 15, 2019 23:57:11 GMT
We successfully generated autosomal genome-wide data and mitochondrial genomes of 10 new individuals from key sites in the Iberian Peninsula ranging from ∼13,000–6,000 calibrated years before present (years cal BP): Late Upper Paleolithic (n = 2), Mesolithic (n = 1), Early Neolithic (EN; n = 4), and Middle Neolithic (n = 3) (Figure 1, Data S1A, and STAR Methods). We furthermore improved the sequencing depth of one Upper Paleolithic individual from Troisième caverne of Goyet (Belgium) dated to ∼15,000 years cal BP and associated with the Magdalenian culture [2]. For each individual, we generated multiple DNA libraries with unique index pairs [10, 11] for next-generation sequencing from ancient DNA (aDNA) extracted from teeth and bones [12] (Data S1B and STAR Methods). These were subsequently enriched using targeted in-solution capture for ∼1240K informative nuclear SNPs [13], and independently for the complete mitogenome [14], and sequenced on Illumina platforms. All libraries contained short DNA fragments (46–65 bp length on average) with post mortem deamination patterns characteristic for aDNA (4%–16% for partial uracil-DNA glycosylase [UDG] and 19%–31% for non-UDG treated libraries at the first base pair position; Data S1C and S1D and STAR Methods). We estimated contamination rates from nuclear DNA in males to be 1.0%–3.4% for final merged libraries using ANGSD method 2 [15] and for mitogenomes in both sexes to be 0.14%–2.20% using ContamMix [16] (Data S1E and S1F and STAR Methods). After quality filtering, we obtained an endogenous DNA content of 1.3%–29.5% on the targeted 1240K SNPs (Data S1C and STAR Methods). We called pseudo-haploid genotypes for each individual (merged libraries) by randomly choosing a single base per site and intersected our data with a set of global modern populations genotyped for ∼1240K nuclear SNP positions [17], including published ancient individuals from [2, 5, 7, 8, 9, 13, 14, 18, 19, 20, 21, 22, 23, 24, 25]. The final dataset from the newly reported individuals contained 19,269–814,072 covered SNPs (Data S1G and STAR Methods). For principal-component analysis (PCA), we intersected our new data and published ancient individuals with a panel of worldwide modern populations genotyped on the Affymetrix Human Origins (HO) array [26].  Figure 1 Geo-chronological Location of Ancient Individuals from the Iberian Peninsula To characterize the genetic differentiation between HG individuals, we calculated genetic distances, defined as 1 – f3, where f3 denotes the f3-outgroup statistics [26], for pairwise comparisons among all published and newly generated HG and visualized the results using multidimensional scaling (MDS) (STAR Methods, Figure 2A and STAR Methods). The hunter-gatherer (HG) individuals form distinguishable clusters on the MDS plot, supported by f4-statistics and clustering analysis (Figure S1 and STAR Methods), which we label in line with Fu et al. [2] as Villabruna, Věstonice, Satsurblia, and Mal’ta clusters, respectively. Henceforth, we present genetic clusters in italics and individuals in normal print. We introduce the GoyetQ2 cluster (based on the highest genomic coverage) representing the Magdalenian-associated individuals Goyet Q-2, Hohle Fels 49, Rigney 1, El Mirón, and Burkhardtshöhle. With the newly generated data, we notice that Iberian HGs form a cline between the GoyetQ2 and Villabruna clusters (Figures 2A and S2). This cline also includes El Mirón (the oldest individual from Iberia), which had previously been considered representing its own El Mirón cluster together with all individuals of the GoyetQ2 cluster (yellow symbols in Figure 2A) [2]. Here, Canes 1 and La Braña 1 (Mesolithic individuals from the Cantabrian region in northern Iberia) are falling closer to the Villabruna cluster, while Chan (northwestern Iberia) and our newly reported individuals from Moita do Sebastião (Portuguese Atlantic coast) and Balma Guilanyà (Pre-Pyrenean region, northeastern Iberia) have more affinity with El Mirón, which is in turn closer to the Magdalenian GoyetQ2 cluster.  Figure 2 Genetic Distances between European HGs and Key f4-Statistics This observation is confirmed by f4-statistics of the form f4(GoyetQ2, Villabruna; test, Mbuti), which measures whether a test population shares more genetic drift with Goyet Q-2 than with the Villabruna individual. Three Iberian HGs (Chan, Moita do Sebastião, and El Mirón), as well as Hohle Fels 49 and the 35,000-year-old Goyet Q116-1, show significantly positive f4-values, indicating that these individuals shared more ancestry with Goyet Q-2 than with Villabruna (Figure 2B). This heterogeneity in Iberian HGs cannot be explained by genetic drift alone (against which this type of F-statistics is robust) but only by admixture between two sources related to Goyet Q-2 and Villabruna, respectively. We visualize this admixture cline using contrasting f3-outgroup statistics of the form f3(GoyetQ2; test, Mbuti) and f3(Villabruna; test, Mbuti) (Figure 3A). The individuals from the Villabruna cluster deviate from the symmetry line x = y toward the y axis, expectedly, indicating excess genetic drift shared with Villabruna. In contrast, individuals of the GoyetQ2 cluster deviate from the symmetry line x = y toward the x axis, indicating excess genetic drift with Goyet Q-2. Iberian HGs fall between the two clusters, which is inconsistent with them forming a clade with either group, but can only be explained by admixture. Here, Iberian HG Canes 1 and La Braña 1 share more Villabruna-like ancestry while El Mirón, Moita do Sebastião, and Chan share more Goyet Q-2-like ancestry.  Figure 3 Key f3-Outgroup Tests, f4-Statistics, and qpAdm Results To further confirm the potential admixture of El Mirón, we used f4(GoyetQ2 cluster, El Mirón; Villabruna, Mbuti) to test if Magdalenian-associated individuals were cladal with El Mirón. Here, we obtained significantly negative Z scores for Hohle Fels 49, Goyet Q-2, and Burkhardtshöhle. Among these, Goyet Q-2 has the highest data quality and most negative Z score and thus represents the best proxy for the non-Villabruna-like ancestry proportion in individuals such as El Mirón (Z = −6.82) (Data S1H). Based on this observation, we used the test f4(Goyet Q-2, GoyetQ2 cluster; Villabruna, Mbuti), for which El Mirón is significantly negative (Figure 3B and Data S1H), confirming shared ancestry with Villabruna. To show that the affinity of El Mirón with the Villabruna individual cannot be explained by El Mirón representing a basal split from Villabruna and the other GoyetQ2 individuals, we used the test f4(GoyetQ2 cluster, Villabruna; El Mirón, Mbuti). Here, all individuals of the GoyetQ2 cluster are significantly positive, indicating that this cluster does not represent a sister branch of Villabruna and that El Mirón is not an outgroup to both the Villabruna and GoyetQ2 clusters (Figure 3C and Data S1H). The mixed ancestry of El Mirón could also explain its reduced affinity to Goyet Q116-1 when compared to the younger GoyetQ2 cluster in the test f4(GoyetQ2 cluster, Goyet Q-2; Goyet Q116-1, Mbuti) (Data S1H). Having established two potential Paleolithic source populations surviving in Iberia from ∼19,000 years cal BP onward, we used the admixture modeling programs qpWave and qpAdm (Figure 3D and STAR Methods) to explore the ancestry of all Iberian HGs. We used Villabruna and Goyet Q-2 as ultimate sources to model the dual ancestry in European HGs relative to outgroups that can distinguish these two sources from shared deeper ancestries (STAR Methods). Our two-source admixture model provides a good fit for the genetic profiles of most European HGs and is consistent with the cline between Villabruna- and Goyet Q-2-like ancestries described above (Figure 3D and STAR Methods). Here, Villabruna-like ancestry is the dominant component (69.8% ± 4.3% − 100%) in individuals of the Villabruna cluster, and Goyet Q-2-like ancestry the dominant component (61.9% ± 6.3% − 94.3% ± 5.7%) in GoyetQ2 cluster individuals (Figure 3D and Data S1I). These results underline the power of our outgroups and choice of proxies to differentiate Goyet Q-2- from Villabruna-like ancestry within our test individuals (STAR Methods; see Figure S3A for a replication with more proximal sources El Mirón and Loschbour). Congruent with the pattern observed in MDS (Figure 2A), clustering analysis (Figure S1), PCA (Figure S2), F-statistic-based tests (Figure 2B), and the biplot of f3-outgroup tests (Figure 3B), the two-source admixture model assigns a higher proportion of Goyet Q-2-like ancestry to Iberian HGs (ranging from 23.7% to 75.3%) than to contemporaneous Western HG (WHG) outside of Iberia. In fact, Balma Guilanyà, La Braña 1, and Canes 1 show elevated Villabruna admixture proportions but still higher Goyet Q-2 proportions than non-Iberian HGs (Figure 3D and Data S1I). We notice an additional contribution of Villabruna-like ancestry in the 12,000-year-old Balma Guilanyà individual from northeastern Iberia. Villabruna-like ancestry becomes even stronger during the Mesolithic in the Cantabrian region (La Braña 1 and Canes 1), suggesting extra HG flux into north/northeastern Iberia, which must have had a higher impact in this region. We were able to track a correlation between increasing Villabruna-like ancestry and time in this region (Figure S3A), while Mesolithic HGs outside this region (Chan and Moita do Sebãstiao) retain more GoyetQ2 ancestry and do not fit this pattern (Figures S3A and S3C). Interestingly, we find no traces of African ancestry (Figure S3B and STAR Methods). |
|
|
|
Post by Admin on Mar 16, 2019 18:27:03 GMT
Phenotypic traits analysis We extracted a list of 36 SNPs of functional importance or related to known phenotypic traits [22] (e.g., lactase persistence, pigmentation, eye colors) (Figure S4) and calculated the genotype likelihood based on the number of reads (using a quality filter q30) for each specific position to determinate the presence of the ancestral or derived alleles [22].  We interrogated different SNPs positions on the gene OCA2 related to light eye color. We obtain the ancestral allele (rs12913832, 3 reads) in individuals CHA001 and ELT002, and heterozygous allele calls for ELT006, suggesting dark color eyes for all of them. The SNP coverage was not sufficient to reliable type the remaining individuals. Another allele from the same gene (rs1800404) related to eye pigmentation could support darker pigmentation in ELT006 than in ELT002. We also checked different SNPs positions in the gene SLC45A2. We obtain the ancestral alleles (rs1426654, 2 reads) in BAL0051 and (rs16891982, 4 reads) in CHA001 which suggest a darker skin color than ELT002 and ELT006, who are heterozygous or homozygous for the derived allele. The coverage in the other individuals is very low to allow comparisons. The allele rs3827760 of the EDAR gene, related to straight and thick hair, is ancestral in all individuals, albeit with variable coverage (CHA001, 4 reads; CHA003, 3 reads; ELT002, 14 reads and; ELT006, 8 reads). Also, as reported before for pre-farming and Neolithic individuals [13], none of our newly typed individuals show evidence for Lactase persistence. Individuals from the Neolithic times (ELT002, ELT006 and FUC003) show different combinations of derived and ancestral alleles of the gene rs174546, which is related to the capacity of regulation of the production of long-chain polyunsaturated fatty acids (FADS1/FADS2). Results are shown in Figure S4. Mitochondrial and Y chromosome haplogroups Using an in-house mtDNA capture assay, we could recover complete mitochondrial genomes from individuals CHA002, CHA003, CHA004, ELT002, ELT006, and FUC003. The coverage of the mtDNA genome for the rest or the samples ranges from 88.86%–99.99% (Data S1D). We used samtools v1.3.1 to extract reads from mitocapture data [31] and mapped them to the rCRS and called the consensus sequences using Geneious R8.1.974 [36]. We downloaded these consensus sequences in fasta format and they were used to determinate mitochondrial haplotypes using Haplogrep 2 [34] (Data S1F). Iberian HG individuals from Balma Guilanyà and Moita do Sebastião belong to haplogroup U, together with the two MN individuals CHA004 and ELT006 (Data S1F). Individual BAL003 could be assigned to U2′3′4’7’8’9’, also found in the Paglicci 108 (∼27,000 years cal BP, Italy), Rigney 1 (∼15,500 years cal BP, France) [2], and Grotta d’Oriente C HG (∼14,000 years cal BP, Italy) [19]. Individual BAL0051 belongs to U5b2a, also found in Neolithic Scotland [9]. Moita do Sebastião (CMS001) carries haplogroup U5b1, which was reported from MN, Bell Beaker and Middle Bronze Age individuals from Portugal and Spain [7, 9], and in the Ranchot 88 HG (∼10,000 years cal BP, France) [2].
Early Neolithic individuals from Cueva de Chaves do not carry U haplogroups. Individual CHA001 could be assigned to haplogroup HV0+195, but was previously reported as K based on PCR-based results [70]. This haplogroup has been reported from MN Ireland [20], as well as MN Scotland and Bell Beaker individuals from England [9]. Individual CHA002 was assigned to K1a2a, which is common in Early Neolithic Iberia, e.g., Cova Bonica [8], Cova de Els Trocs [3] and Cueva del Toro [30], but also in Chalcolithic and Bell Beaker individuals from Iberia and Italy [9, 13]. CHA003 was assigned to K1a3a, so far reported from Neolithic Anatolia [13], Neolithic and Chalcolithic Scotland, and Bell Beaker individuals from Sicily [9]. Middle Neolithic individual CHA004 carried haplogroup U4a2f, found in HG from the Iron Gates, Romania, and Lithuania [14, 19]. The MN individual ELT002 carries haplogroup J1c1b, present in the Körös Early Neolithic [18], Neolithic from Scotland [9], Iberian Late Neolithic-Chalcolithic [7], and Bronze Age from Italy and Germany [6]. ELT006 was assigned to haplogroup U3a1, which has been reported from MN France and Germany [9, 19], and Chalcolithic Iberia [13]. Early Neolithic Fuente Celada carries haplogroup X2b+226, found in MN Hungary [71] and Middle Bronze Age Iberia [7]. X2b was found in Iberian Late Neolithic [5], Chalcolithic [18] and Bell-Beaker individuals [9], Neolithic England [9] and Greece [72], and EN/LN Morocco [25].
BAL0051 could be assigned to haplogroup I1, while BAL003 carries the C1a1a haplogroup. To the limits of our typing resolution, EN/MN individuals CHA001, CHA003, ELT002 and ELT006 share haplogroup I2a1b, which was also reported for Loschbour [73] and Motala HG [13], and other LN and Chalcolithic individuals from Iberia [7, 9], as well as Neolithic Scotland, France, England [9], and Lithuania [14]. Both C1 and I1/ I2 are considered typical European HG lineages prior to the arrival of farming. Interestingly, CHA002 was assigned to haplogroup R1b-M343, which together with an EN individual from Cova de Els Trocs (R1b1a) confirms the presence of R1b in Western Europe prior to the expansion of steppe pastoralists that established a related male lineage in Bronze Age Europe [3, 6, 9, 13, 19]. The geographical vicinity and contemporaneity of these two sites led us to run genomic kinship analysis in order to rule out any first or second degree of relatedness. Early Neolithic individual FUC003 carries the Y haplogroup G2a2a1, commonly found in other EN males from Neolithic Anatolia [13], Starçevo, LBK Hungary [18], Impressa from Croatia and Serbia Neolithic [19] and Czech Neolithic [9], but also in MN Croatia [19] and Chalcolithic Iberia [9].Dual Hunter-Gatherer Genetic Legacy in Iberian Neolithic Individuals During the Neolithic transition ∼7,600 years ago, human expansions reached the Iberian Peninsula relatively swiftly via expanding early farmers from western Anatolia [3, 4, 5]. The rapid expansion of EN individuals associated with farming practices across Europe resulted in a relatively low genetic variability in the reported Neolithic genomes, which makes it difficult to distinguish between the Mediterranean and Danubian routes of expansion of Neolithic lifeways [27, 28]. However, Olalde et al. [9] noted subtle regional differences between WHG individuals and used the proportion of HG ancestry from La Braña 1 in Neolithic Iberians to trace the expansion from southwestern Europe along the Atlantic coast to Britain. This movement corresponds well with the megalithic burial practices of these regions observed in the archaeological record [29, 30]. Under the assumption that these proportions reflect one, or potentially more, local admixture events along the routes of expansion, it is thus possible to distinguish Neolithic groups by their varying autochthonous HG signatures [18]. Given the presence of two ancestral lineages in Iberian HGs, we thus explored this potential genetic legacy in our newly generated EN and Middle Neolithic (MN) individuals. We first used PCA to assess the genetic affinities qualitatively (Figure 4A). Here, the new Neolithic Iberian individuals spread along a cline from Neolithic Anatolia to WHG, on which the new individuals cluster with contemporaneous Iberian Neolithic individuals [5, 7, 8, 13, 24, 25]. As shown before, MN individuals are shifted toward WHG individuals [3], including the newly reported MN individuals from Cova de Els Trocs and Cueva de Chaves.  Figure 4 PCA Results and qpAdm Admixture Models Using qpAdm models consistent with those above (Figures 3D and S3), we aimed to trace and quantify the proportion of Goyet Q-2- and Villabruna-like HG ancestry in EN and MN groups from Iberia and western/central Europe as a mixture of three ancestral sources: Anatolian Neolithic, Goyet Q-2, and Villabruna, respectively. We show that EN Iberians shared a higher proportion of Goyet Q-2-like ancestry than EN individuals from outside Iberia (Figure 4B and Data S1J). Goyet Q-2-like ancestry is higher in EN from southern Iberia (Andalusia), suggesting additional admixture with local Iberian HGs, who carried mixed Upper Paleolithic ancestry. Goyet Q-2 ancestry is continuously detectable in all Iberian MN individuals, including broadly contemporaneous individuals from Scotland, Wales, Ireland, and France, but not in Neolithic England, for which the qpAdm model with three sources fails (p = 6.91e−05) in favor of two sources, despite being poorly supported (p = 0.001) (Data S1J). Goyet Q-2-like ancestry is, however, highest in all Iberian MN (except Chaves MN) when compared with other MN populations that share a similar overall amount of HG ancestry (Figures 4B and 4C). We note the presence of Goyet Q-2 ancestry in MN Trocs, where this ancestry was not observed during the EN, but importantly also in MN individuals from France and Globular Amphora from Poland. Olalde and colleagues [9] reported an elevated signal of La Braña 1 ancestry in Neolithic individuals from Wales and England (using KO1 HG from Hungary and Anatolian Neolithic as the other two sources) and thus argued for an Iberian contribution to the Neolithic in Britain [9]. We replicated these findings by using similar sources (El Mirón instead of Goyet Q-2; Data S1J), showing that these results are sensitive to the source populations used. However, our models with Goyet Q-2 as ultimate source highlight not only the admixed nature of La Braña 1 and El Mirón, but also that Goyet Q-2-like ancestry in MN individuals outside Iberia hints at Iberia as one possible source, but not the exclusive source, of the Neolithic in Britain. Further sampling from regions in today’s France, the Netherlands, Belgium, Luxembourg, and Germany is needed to answer this question. Conclusions Our results highlight the unique genetic structure observed in Iberian HG individuals, which results from admixture of individuals related to the GoyetQ2 and Villabruna clusters. This suggests a survival of two lineages of Late Pleistocene ancestry in Holocene western Europe, in particular the Iberian Peninsula, whereas HG ancestry in most other regions was largely replaced by Villabruna-like ancestry. With an age estimate of ∼18,700 years cal BP for the El Mirón individual, the oldest representative of this mixed ancestry, the timing of this admixture suggests an early connection (terminus ante quem) between putative ancestries from different LGM refugia. It is possible that Goyet Q-2 ancestry could have existed in Iberia in unadmixed form, where it was complemented by Villabruna ancestry as early as ∼18,700 years ago. Alternatively, both Magdalenian-associated Goyet Q-2 and Villabruna ancestries originated in regions outside Iberia and arrived in Iberia independently, where both lineages admixed, or had already existed in admixed form outside Iberia. Interestingly, the dual Upper Paleolithic ancestry was also found in EN individuals from the Iberian Peninsula, supporting the hypothesis of additional local admixture with resident HGs in Iberia during the time of the Mesolithic-Neolithic transition. Published: March 14, 2019 DOI:https://doi.org/10.1016/j.cub.2019.02.006 |
|
|
|
Post by Admin on Mar 17, 2019 22:23:13 GMT
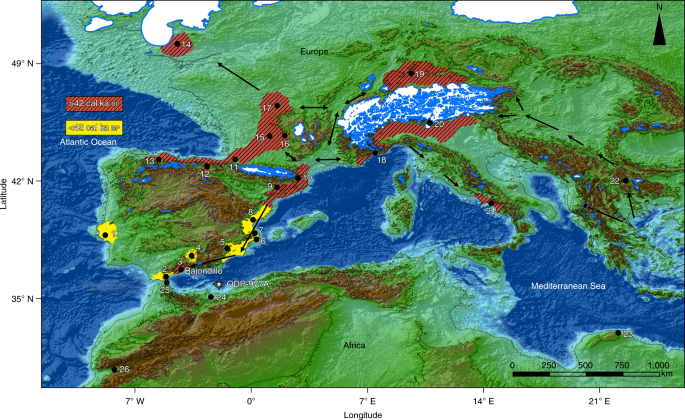 Fig. 1: Selected archaeological sites in western Europe and North Africa. Great effort has been devoted to framing the spatiotemporal fea-tures of Neanderthal replacement, as this may help to resolve the processes that triggered population and technological turnovers6. This includes determining any directionality of technological and population changes that, according to genetic and archaeological data, become recurrent events in western Europe from the Late Pleistocene onwards, generally exhibiting an east to west trend. Mousterian technocomplex replacement by Aurignacian popula-tions is postulated to be one such east-to-west population turn-over1. This same directionality is documented for the Gravettian technocomplex that replaced the Aurignacian7, the genetically and culturally documented Magdalenian–Azilian transition in western Europe at the beginning of the Bölling–Alleröd interstadial8, and replacements within the Epigravettian cultures of southern Europe.Among European east–west population turnovers, the Mousterian–Aurignacian transition has perhaps received the most attention. This is because it is associated with the putative extinction of Neanderthals, given that Aurignacian technocomplex elements have now been securely associated with AMHs1. In comparing the early stages of the Aurignacian dispersal (Fig. 1) with subsequent transitions, two spatiotemporal anomalies emerge. The first is recorded on the Italian Peninsula, where populations manufacturing the Uluzzian industry (for some authors, a develop-ment rooted in the Mousterian lithic tradition9) seemingly prevented the expansion of the early Aurignacian10. The second anomaly is documented in mid-southern Iberia, where the Aurignacian expan-sion is postulated to have been delayed to the point of failure11. The proposal that the Middle Palaeolithic technocomplex extended to the end of Marine Isotope Stage 3 (Gorham’s Cave, Gibraltar: ~32.5 calibrated thousand years before the present (cal ka BP11) has lent weight to the hypothesis that the EUP reached southern Iberia at a comparatively late date. This reinforced the validity of a particu-lar version of the east-to-west wave-of-advance mode that set apart the ‘Iberian South’ from the rest of the Peninsula. 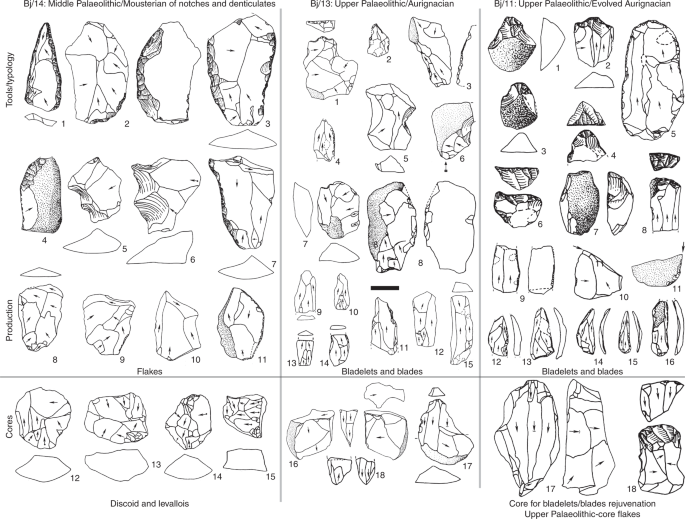 Fig. 2: Representative lithic industries from Bajondillo site archaeological levels Bj/14–Bj/11. To reliably pin down the Neanderthal–AMH transition in southern Iberia, 17 new dates restricted to levels covering the transition of the Mousterian to the Aurignacian at the site of Bajondillo in the Bay of Málaga (southern Spain) (Fig. 1, Supplementary Figs. 1–3 and Supplementary Table 1) have been integrated with previous radio-metric dates13, using a Bayesian approach. The Middle Palaeolithic at Bajondillo lasts for ~120 kyr14, as represented by 6 archaeological levels (that is, Bj/19–Bj/14) (Supplementary Fig. 1), the last of which features a Denticulate Mousterian (~50–46 cal ka)13 (Fig. 2). The evidence that allows us to identify Bj/13 as Proto-Aurignacian or Early Aurignacian results from a combination of chronological and stratigraphic data, complemented by historiographic and tech-nological data (Fig. 2 and Supplementary Fig. 4). Stratigraphically, Bj/13 is firmly set above a well-defined Middle Palaeolithic pack-age and overlaid by another Aurignacian level (Bj/11) lying below a Gravettian deposit (Bj/10). The stratigraphic package is well structured in sedimentological and micromorphological terms, and shows no inconsistency in the seriation of absolute dates indepen-dent of the long- or short-lived nature of the samples (see below). The range of dates obtained from the Bayesian model appears to be fully consistent with our hypothesis of the Bj/13 Aurignacian (43.4–40.0 cal ka BP) lying chronologically between Heinrich events 5 (50–47 cal ka BP) and 4 (40.2–38.3 cal ka BP). However, techno-logically, level Bj/11 (~37.6–32.4 cal ka BP; Fig. 2 and Supplementary Table 1) corresponds to an Evolved Aurignacian with blade and bladelet technologies and the characteristic tools of this techno-complex. Hearths also testify to differences between the Middle Palaeolithic and EUP. Hearths from Bj/16 and Bj/15 evidence an exclusive use of grasses (Gramineae) as fuel, with temperatures never reaching 500 °C, whereas those from EUP levels (that is, Bj/12 onwards) exhibit a more efficient pyrotechnology, with frequent use of wood, and temperatures well in excess of 550 °C20.At Bajondillo, all six of the new accelerator mass spectrom-etry (AMS) dates for the Mousterian from Bj/14 derive from short-lived (< 5 years) shells of marine and terrestrial molluscs, and range between > 50 and 46 cal ka BP, the youngest ages being based on a Bayesian model (Supplementary Table 1; the pulmo-nate samples derive from a garden snail taxon (Otala species) that feeds on fresh vegetal matter and thus does not exhibit the ‘old-shell effect’). These dates calibrate this level to during or immedi-ately after Heinrich event 5 (Fig. 3). The date obtained on a mussel from the latest Middle Palaeolithic level at the nearby Abrigo 3 site, 20 km away from Bajondillo Cave on the Bay of Málaga, falls squarely within this range of dates (that is, 50–43.1 cal ka BP21; Supplementary Fig. 2b). 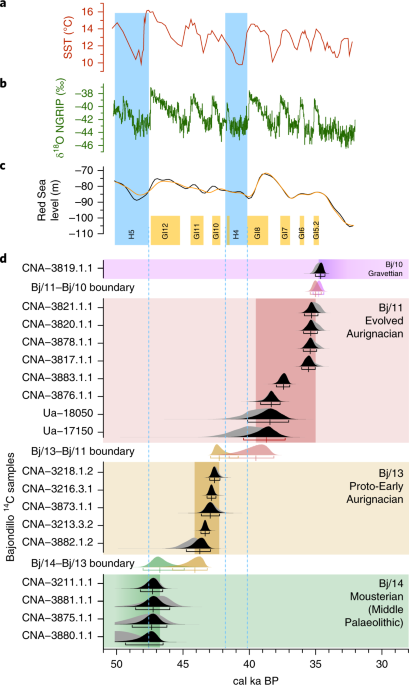 Fig. 3: Comparison between the chronologies of the different archaeological levels at Bajondillo Cave and a selection of palaeoenvironmental proxies. The oldest of the six AMS dates from Bj/13 (EUP) come from long-lived taxa (sample charcoal/Ua-18270) (Supplementary Table 1), which, although coherent among these taxa, may yield ages older than the deposit from which they were retrieved. Samples for Bj/13 that derive from short-lived (< 5 years) mussels and a terrestrial snail (Otala species) provide a Bayesian median age set between 42.5 and 41.2 cal ka BP (Fig. 3 and Supplementary Table 1). The taphonomic features (that is, fragmentation and thermoalteration) of all marine shells from Bj/13 document shellfish processing contemporane-ous with the deposit, reinforcing the autochthonous nature of the radiocarbon-dated samples, thus their reliability as chrono-indica-tors. The dates available from Bj/14 and Bj/13 (which were derived from the middles of the layers, away from their common bound-ary) show a potential discontinuity of ~2 kyr between these levels, one of which requires further research to be confirmed (Fig. 3 and Supplementary Table 1). The dates from Bj/13 coincide with others from early Aurignacian sites in Europe (Fig. 1), such as the Italian rock shelter of Riparo Mochi, as well as sites from Central Europe (~43-42.5 cal ka)23 and (~43.5 cal ka)24. Additionally, the most recent Middle Palaeolithic/Upper Palaeolithic chronological review from the Cantabrian area (northern Spain)25 and Central Spain26 yields essentially identical dates to those obtained at Bajondillo for the Middle Palaeolithic–Upper Palaeolithic transition. In North Africa, Upper Palaeolithic/Later Stone appear in Cyrenaica after ~43 ka (Haua Fteah), but only after 30 ka in the Maghreb (Grotte des Pigeons) (Fig. 1)8,12,17,27–34. The new Bajondillo dates are crucial for several reasons. First, they confirm the presence of a chronologically early Aurignacian in southern Iberia at ~43 cal ka BP that now shows the first appear-ance of the EUP to be an essentially synchronous event throughout Europe (Fig. 3 and Supplementary Fig. 3). This suggests that the dis-persal of AMHs was much faster than hitherto postulated, and the expansion of the earliest Aurignacians in Europe is now increased westwards by > 1,000 km. These dates thus call into question both the gradual ‘wave-of-advance’ and the ‘Ebro frontier’ models. They also provide reference points for the attribution of early art work in Iberian caves—something that remains highly controversial35. The dates suggest either extremely high mobility of early Aurignacians or well-developed networks of interchange. For early AMHs, rapid dispersal was seemingly only possible over essentially ‘empty’ terri-tories (that is, either completely depopulated areas, as some studies suggest for southern Iberia at the time7, or areas featuring severely depleted human populations).The Aurignacian early expansion took place between Heinrich events 5 and 4 (Fig. 3 and Supplementary Fig. 3), when cold and steppic conditions prevailed throughout most of western Europe, including the Iberian hinterland36. In light of this, it might be no coincidence that a prevalence of European EUP sites has been reported along shores or neighbouring lowlands where milder con-ditions prevailed and more productive environments, in terms of living resources, existed (Fig. 1 and Supplementary Fig. 2c). The Aurignacian from Bajondillo Cave conforms with this pattern and is not an isolated case in Iberia, where newly recorded Aurignacian sites south of the Ebro river, such as Foradada16, Cendres17 and Pego do Diabo12, are all located on the present-day coast or its adjoin-ing lowlands (Fig. 1), nor in Italy (Riparo Mochi and Serino; see Supplementary Information). Coasts and coastal lowlands as instrumental for human dispersal and colonizing events are not a phenomenon restricted to the European Aurignacian, since data emerging from southern Arabia37, Australia38,39 and South America40 all point in the same direction. In eastern and southern Iberia, trav-elling through coasts and coastal lowlands would have been con-siderably easier than travelling through one of the most rugged mountainous hinterlands in Europe. The enhanced mobility of Aurignacian populations can be inferred from their swift spread over western Europe from Glacial Interstadial 12 onwards, thus hinting at dispersals taking place across territories that were easy to traverse (Supplementary Fig. 2a,b)1.The onset of the Aurignacian cannot be detached from the demise of the Neanderthals. Inferences about the mobility and settlement patterns of southern Iberian Neanderthals are compli-cated due to the restricted number of Mousterian sites, as well as the prevalence of low sea-level stands during this period, which means that an undetermined, yet probably substantial, fraction of the evidence presently lies underwater. From this perspective Bajondillo Cave is of great relevance since, due to its topogra-phy, it was at all times located on or very close to the shore (that is, not more than 5 km distant; Supplementary Fig. 2a), but never flooded. The data from Bajondillo Cave and other coastal Iberian locations41 reveal that stasis, as exemplified by around 120 kyr14 of Neanderthal occupation with no clear traces of technological devel-opments, prevailed during the Iberian Mousterian. Nature Ecology & Evolutionvolume 3, pages207–212 (2019) |
|

