|
|
Post by Admin on Aug 7, 2018 18:31:58 GMT
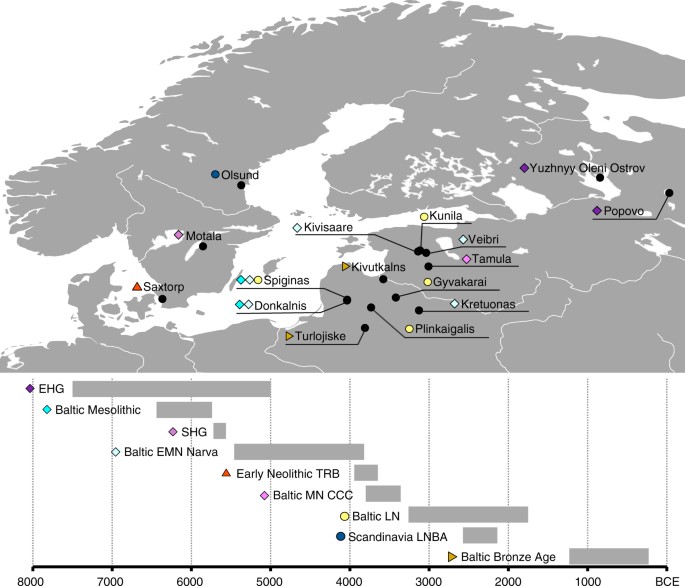 Fig. 1 Sampling locations and dating of 38 ancient Northern European samples introduced in this study. Chronology based on calibrated radiocarbon dates or relative dating, see Supplementary Note 2. Map generated with QGIS 2.18.2 (http://www.qgis.org/) using the Natural Earth data set (http://www.naturalearthdata.com) for the basemap The skeletal remains studied here were recovered from 25 archaeological sites in the territory of modern Lithuania, Latvia, Estonia, Archangelsk Oblast and Karelia (north-western Russia) and Sweden dating from around 7500 to 200 calBCE (Fig. 1, Supplementary Note 2, Supplementary Data 1). In total, we analyzed DNA from 106 human remains. A total of 41 samples with good DNA preservation were selected for deeper shotgun sequencing or SNP capture (Supplementary Data 1). In the latter case, we enriched samples for a panel of around 1.24 million single nucleotide polymorphisms (SNPs) via in-solution capture4,24. After quality control, genome-wide data from 38 individuals, with an average coverage of 0.02–8.8-fold on targeted SNPs, were included in further analysis. We supplemented our dataset of ancient Eastern Baltic samples with recently published data from 13 individuals spanning the Mesolithic to Late Neolithic in Latvia21 and Estonia22, and merged data from identical individuals where they overlapped with the latter. We present the first data from the southernmost region of the Eastern Baltic (the territory of modern-day Lithuania), the Early Neolithic of Estonia and the Eastern Baltic Bronze Age.  From southern Sweden, we analyzed one farmer (3950–3650 calBCE) from the EN TRB, the earliest agricultural population in Scandinavia for which there exists no genetic data to date. One sample from northern Sweden (Scandinavia LNBA Olsund, ca. 2570–2140 calBCE) dates to the Late Neolithic but was found without associated archaeological assemblages. The data were analyzed in context with published data from the Scandinavian Middle Neolithic to Bronze Age3,19 as well as other ancient and modern genome-wide datasets described below. Affinities of northern Mesolithic hunter-gatherers To gain an overview of the broad genetic affinities of our samples, we projected all 38 ancient genome-wide datasets, as well as previously published ancient samples4,6,19,21,22, using a principal component analysis (PCA), constructed from 1007 modern individuals from a diverse set of West Eurasian contemporary populations, and used the same individuals to investigate model-based clustering using ADMIXTURE. We see that the Mesolithic foragers of Northern Europe fall into three distinct clusters, associated with EHG, SHG and WHG, respectively, as evidenced by their position on the PCA (Fig. 2a), similar composition of ancestral genetic clusters in ADMIXTURE analysis (Fig. 2b, Supplementary Fig. 4) and in sharing most genetic drift since divergence from Africa as shown by outgroup f3-statistics (Supplementary Fig. 1). Based on these results, our Mesolithic Russian foragers fall within the EHG cluster formed by previously published samples4 and are grouped as EHG in subsequent analyses. |
|
|
|
Post by Admin on Aug 9, 2018 18:27:03 GMT
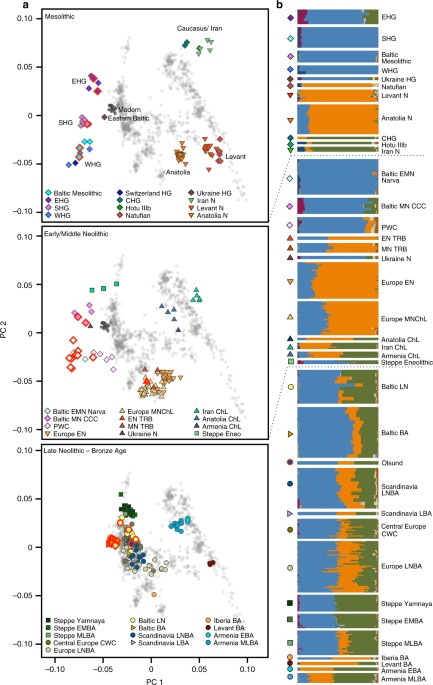 Fig. 2 PCA and ADMIXTURE analysis reflecting three time periods in Northern European prehistory. a Principal components analysis of 1012 present-day West Eurasians (grey points, modern Baltic populations in dark grey) with 294 projected published ancient and 38 ancient North European samples introduced in this study (marked with a red outline). Population labels of modern West Eurasians are given in Supplementary Fig. 7 and a zoomed-in version of the European Late Neolithic and Bronze Age samples is provided in Supplementary Fig. 8. b Ancestral components in ancient individuals estimated by ADMIXTURE (k = 11) PCA and ADMIXTURE analysis reflecting three time periods in Northern European prehistory. a Principal components analysis of 1012 present-day West Eurasians (grey points, modern Baltic populations in dark grey) with 294 projected published ancient and 38 ancient North European samples introduced in this study (marked with a red outline). Population labels of modern West Eurasians are given in Supplementary Fig. 7 and a zoomed-in version of the European Late Neolithic and Bronze Age samples is provided in Supplementary Fig. 8. b Ancestral components in ancient individuals estimated by ADMIXTURE (k = 11) ADMIXTURE shows that EHG carry a genetic component (green component in Fig. 2b) that is maximized in hunter-gatherers from the Caucasus (CHG) and shared with Neolithic farmers from Iran and Steppe populations from the Bronze Age, suggesting some common ancestry for these populations, consistent with previous results21. Despite their geographical vicinity to EHG, the two Eastern Baltic individuals associated with the Mesolithic Kunda Culture show a very close affinity to WHG in all our analyses (Fig. 2, Supplementary Figs. 1 and 2), with a significant contribution from ANE, as revealed by negative admixture f3 results involving a Palaeolithic hunter-gatherer from Switzerland, most closely related to WHG, and populations containing ANE ancestry (Supplementary Table 1). This is additionally confirmed by D-statistics of the form D(Baltic Mesolithic, WHG; X, Mbuti) for populations X with ANE ancestry, which are significant and among the highest in EHG (Z = 6.2; Supplementary Table 2). Using the qpWave/qpAdm framework, we modelled the Baltic Mesolithic hunter-gatherers as a two-way mixture between EHG and WHG (Fig. 3), which reveals a difference in mixture proportions between the more northern individuals from the Latvian site21 (65–76% WHG with 24–35% EHG; Supplementary Table 3) and the samples from the Lithuanian sites to the south (88–100% WHG with 0–12% EHG; Supplementary Table 3). |
|
|
|
Post by Admin on Aug 11, 2018 18:43:24 GMT
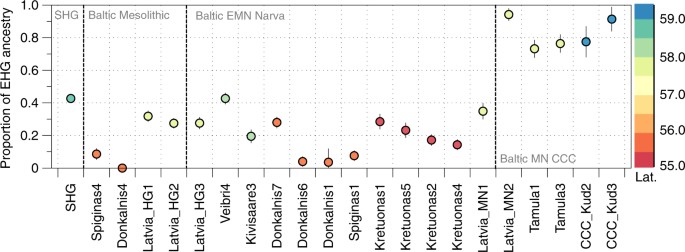 Fig. 3 EHG ancestry in SHG and Eastern Baltic hunter-gatherers. Ancestry proportions were estimated with qpAdm, and standard errors are shown as vertical lines. Colours indicate latitude of the site at which the individuals were excavated SHG appear intermediate between WHG/Baltic Mesolithic and EHG in PCA space, have increased shared genetic drift with both shown in outgroup f3-statistics (Supplementary Fig. 3) and the statistic D(SHG,WHG; EHG, Mbuti) is strongly significant for excess allele sharing of SHG and EHG (Z = 7.3; Supplementary Table 4). Using qpAdm, we confirm the previously published result of SHG being formed by admixture of WHG and EHG6 (57 ± 2% WHG with 43 ± 2% EHG; p = 0.19; Supplementary Table 3). Both EHG and SHG share a non-negligible component in ADMIXTURE analysis that is maximized in some modern Native American populations which points towards ANE ancestry, as represented by the MA1 and AG3 samples from Palaeolithic Siberia12 (maroon component in Fig. 2a, Supplementary Fig. 4). Indeed, D-statistics show that EHG and SHG share significantly more alleles with MA1 and AG3 than WHG (Supplementary Tables 2 and 4). Additionally, mtDNA haplogroups found among EHG point towards an eastern influence (Table 1): R1b in UzOO77 (this individual had previously been assigned to haplogroup H58) was also found in the Palaeolithic Siberian AG35 and a haplogroup within the C1 clade, which appears today in highest frequencies in north-east Asia and the Americas, was described in several samples from Yuzhnyy Oleni Ostrov25,26. It was shown that some SHG carry the derived variant of the EDAR allele, which affects hair thickness and tooth morphology among other things, and which is found today in high frequency in East Asians and Native Americans4. Dynamic forager networks in the Eastern Baltic Neolithic Similarly to the Baltic Mesolithic, the later Eastern Baltic Neolithic hunter-gatherers of the Narva culture exhibit varying proportions of EHG (0–46%) and WHG (54–100%) ancestry (Fig. 3 and Supplementary Table 3). In principle, it is possible that such a pattern is not the result of admixture but a signal of a long-standing geographic cline. However, in this case, it appears to be more compatible with recent admixture between differentially WHG- and EHG-related groups, as we see varying ancestry proportions even within contemporary individuals from the same site or closely located sites (Fig. 3). D-statistics of the form D(Baltic EMN Narva, WHG; X, Mbuti) do not show evidence of admixture with the contemporaneous European farming populations that were related to the Anatolian Neolithic (Supplementary Tables 2 and 5). The later individuals attributed to the Baltic MN CCC exhibit a significantly higher affinity to EHG with the ancestry proportion estimated at 68–99% EHG and 1–32% WHG (Fig. 3, Supplementary Table 3). Similar to other European Mesolithic hunter-gatherers, our Baltic foragers carry a high frequency of the derived HERC2 allele which codes for light iris colour, and like SHG and EHG they already possess an increased frequency of the derived alleles for SLC45A2 and SLC24A5, coding for lighter skin colour (Supplementary Table 6). The male individuals carry Y-chromosomal haplogroups of the I and R1 clades (Table 1, Supplementary Note 3). Y-haplogroup I has been most commonly found among WHG and SHG1,5 while R1 is found in EHG2 and other published Eastern Baltic21,22 and Romanian hunter-gatherers27.  One Narva individual, Spiginas128, dated to ca. 4440–4240 calBCE, belongs to a mitochondrial haplogroup of the H branch, normally associated with the Neolithic expansion into Europe, but shows no evidence of Neolithic farmer ancestry on the nuclear level suggesting that this haplogroup might have been present already in foraging groups (Table 1, Supplementary Table 5). In addition to haplogroup H, the maternal lineages seen in Eastern Baltic samples (n = 35; Supplementary Fig. 5) encompass all of the major haplogroups identified in complete mtDNA genomes from Holocene Scandinavian and Western European hunter-gatherers (n = 21:U2, U5a, U5b)29, as well as haplogroup U4 which has been found in high frequency in Mesolithic foragers from Russia25. We also find mtDNA branch K1, a subclade of U8, in one Baltic Mesolithic forager, adding to the mounting evidence that this lineage was present at low frequency among European hunter-gatherers before the arrival of agriculturalists16,27. |
|
|
|
Post by Admin on Aug 13, 2018 18:35:25 GMT
 Early farming in Sweden coincides with a shift in ancestry In contrast to the Eastern Baltic, we see clear evidence for the genetic impact of the Neolithic expansion already around 4000 calBCE in southern Sweden. The individual associated with the EN TRB culture clusters with Middle and Early Neolithic farmers from Europe on the PCA (Fig. 2a) and in the ADMIXTURE analysis exhibits the component maximized in Levantine and West Anatolian early farmers (orange component in Fig. 2b). D-statistics show increased allele sharing of EN TRB with hunter-gatherer populations in comparison to the preceding farmers of the Linearbandkeramik (LBK) culture and no difference to the contemporaneous Middle Neolithic and Chalcolithic (MNChL) farmers of Central Europe (Supplementary Table 4), paralleling the previously described resurgence of WHG ancestry during the European Middle Neolithic2,27. Different models of EN TRB as a linear combination of LBK with SHG, WHG or Baltic hunter-gatherers favour the latter two groups, while SHG is rejected as a source (Supplementary Table 3). The previously published succeeding farmers of the Middle Neolithic (MN) TRB culture in West Sweden19 appear as directly descended from the EN TRB, with no significant positive results for D(MN TRB, EN TRB; X, Mbuti) (Supplementary Table 4). The PWC individuals, who were contemporaneous to the MN TRB but relied mainly on marine resources, appear intermediate between SHG and Middle Neolithic farming cultures on the PCA (Fig. 2a). Indeed, the statistic D(PWC, SHG; X, Mbuti) reaches weak significance when X is MN TRB (Z = 2.94) and a two-way admixture model for PWC involving SHG and TRB farmers is not rejected by qpWave/qpAdm (74 ± 6% SHG and 26 ± 6% EN TRB; Supplementary Table 3). |
|
|
|
Post by Admin on Aug 15, 2018 18:41:44 GMT
New networks of contact during the LNBA The substantial population movement at the beginning of the 3rd millennium calBCE, during the European LNBA, affected the genetic makeup of Eastern and Central Europe and Scandinavia2,3. It also made its mark in the Eastern Baltic region, as seen in our five samples from Lithuania and Estonia dated to this period and the previously published individuals from the Eastern Baltic region21,22. All Baltic LN individuals (ca. 3200–1750 calBCE) fall in PCA space in the diffuse European LNBA cluster formed by individuals admixed between Early and Middle Bronze Age pastoralists from the Yamnaya culture of the eastern Pontic Steppe and Middle Neolithic European farmers (Fig. 2a). They all carry the genetic component that was introduced into Europe with this pastoralist migration in varying amounts, and the majority also carries the component associated with Anatolian farmers (green and orange, respectively, in Fig. 2b). This genetic impact is furthermore reflected in the uniparental markers where we see novel mitochondrial haplogroups (I, J, T2, W), not found in the preceding foragers, in half of our samples (Supplementary Fig. 5), and I2a Y-chromosomal haplogroups replaced by R1a types (Table 1, Supplementary Note 3). Computing D-statistics for each individual of the form D(Baltic LN, Yamnaya; X, Mbuti), we find that the two individuals from the early phase of the LN (Plinkaigalis242 and Gyvakarai1, dating to ca. 3200–2600 calBCE) form a clade with Yamnaya (Supplementary Table 7), consistent with the absence of the farmer-associated component in ADMIXTURE (Fig. 2b). Younger individuals share more alleles with Anatolian and European farmers (Supplementary Table 7) as also observed in contemporaneous Central European CWC individuals2. The individual Spiginas230, dated to a very late period of the LN (2130–1750 calBCE), stands out in that it shares an excess of alleles with European forager groups when compared to the Yamnaya populations, with the top hits being Switzerland_HG, WHG, Baltic Mesolithic and Baltic EMN Narva (Supplementary Table 7). Fig. 2 This result is the earliest evidence for a continuing pattern: we observe that the increased affinity to Baltic hunter-gatherers remains prevalent in the more recent samples from the Baltic BA (dated between ca. 1000 and 230 calBCE) that cluster together on the PCA in the same space occupied by modern Lithuanians and Estonians, shifted from other Europeans to WHG and Baltic hunter-gatherers (Fig. 2a). The statistic D(Baltic BA, Baltic LN; Baltic EMN Narva, Mbuti) is strongly significant (Z = 14.0; Supplementary Table 2) demonstrating the increase in allele sharing with local hunter-gatherers in the Baltic populations after the Late Neolithic. Replacing Baltic EMN Narva with the contemporaneous northern population of Baltic MN CCC does not yield significant results, suggesting that admixture with this population did not play a large role in the ancestry of our studied Bronze Age individuals. Additionally, D-statistics provide significantly positive results for D(Baltic BA, Baltic LN; X, Mbuti) when X was replaced by various agricultural populations of Europe and the Near East (Supplementary Table 2), which suggests that the formation of the Baltic BA gene pool was not completed by admixture between the Baltic LN population and foragers but involved additional gene-flow from outside the Baltic territory. Archaeological evidence supports that the site Kivutkalns, which is represented by 10 of our individuals, was a major bronze-working centre located on a trade route that opened to the Baltic Sea on the west and led inland following the Daugava river31, where contact to surrounding populations might have been common. The individual from Olsund in north-eastern Sweden was dated to the Late Neolithic, when agriculture had been introduced to the coastal areas of northern Sweden with the Battle Axe Culture, the regional variant of the CWC, while foraging persisted as an important form of subsistence. The remains were found without any associated artefacts, but in close proximity to a site where the assemblage showed a mix between local hunter-gatherer traditions and CWC influence32. On the PCA plot, this sample falls within the European LNBA cluster (Fig. 2a) and similarly to other individuals from this cluster displays the three components derived from WHG, CHG and Neolithic Levant (Fig. 2b). This provides genomic evidence for the influence of both the early Neolithic and LNBA expansions having reached as far as northern Sweden in the 3rd millennium calBCE, either through a northward expansion from southern Scandinavia or across the Baltic Sea by boat or over the frozen sea33. Assemblages similar to the early Battle Axe Culture of Sweden have been found in south-western Finland, across the Bothnian Sea34,35 which could be considered a geographically closer source than southern Scandinavia. |
|

