|
|
Post by Admin on Aug 16, 2018 18:42:27 GMT
Gene-flow into the Eastern Baltic after the Bronze Age Modern Eastern Baltic populations cluster with Baltic BA on the PCA plot and exhibit among all modern populations the highest shared genetic drift with ancient Baltic populations (Supplementary Fig. 2), but show substantial differences to samples from the Bronze Age. The statistic D(Lithuanian, Baltic BA; X, Mbuti) reveals significantly positive results for many modern Near Eastern and Southern European populations (Supplementary Fig. 6a). Limited gene-flow from more south-western neighbouring regions after the Bronze Age is sufficient to explain this pattern, as nearly all modern populations besides Estonians, especially for Central and Western Europe, have a higher amount of farmer ancestry than Lithuanians. 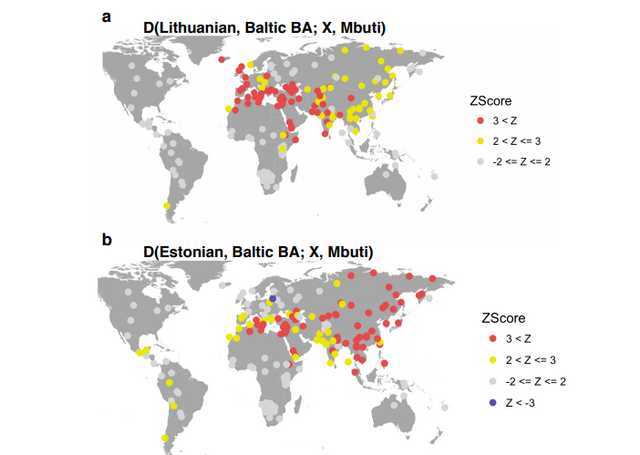 In contrast, the statistic D(Estonian, BA Baltic; X, Mbuti) gives significant positive hits for East Asian and Siberian populations (Supplementary Fig. 6b). None of our male Bronze Age individuals carry Y-haplogroup N, which is found in modern Europeans in highest frequencies in Finland and the Baltic states36. Instead, we observe a high frequency of R1a Y-haplogroups. |
|
|
|
Post by Admin on Aug 18, 2018 18:26:55 GMT
Discussion Our analyses support a dynamic population history of the Baltic Sea region, where populations did not remain in ‘genetic stasis’ despite the late adoption of agrarian subsistence strategies when compared to the rest of Europe. The Mesolithic SHG excavated at Motala, Sweden, owe their genetic signature to an admixture of WHG and EHG and similarly to the latter carry substantial ANE ancestry. In contrast, the two Eastern Baltic Mesolithic Kunda individuals, who predate the SHG, carry comparatively low proportions of ANE ancestry, indicating that this ancestry was never widespread to the southeast of the Baltic Sea and likely reached Scandinavia without traversing the Eastern Baltic. This provides indirect support to the archaeological evidence that Scandinavia was settled by two routes13, suggesting a scenario in which the ANE-related ancestry was brought into Scandinavia with a movement of people via a north-eastern coastal route, where they admixed with a WHG-like population that derived from a migration over the land-bridge that connected Denmark and southern Sweden at the time. This scenario is also supported by the finding that three Mesolithic hunter-gatherers excavated at the coast of Norway carry a higher proportion of EHG ancestry compared to the individuals from inland Sweden37. In southern Scandinavia, the sequence of events resembles that seen in Central Europe, albeit several millennia later, in that the earliest agriculture in the region coincides with the appearance of people related to the Anatolian and European Neolithic. However, similar to Middle Neolithic Central and Southern European populations, early Scandinavian farmers are already strongly admixed with hunter-gatherer groups. Without the knowledge of the genetic substrate in Mesolithic southern Scandinavia, these results are consistent with different scenarios; e.g. a movement of a Central European population into southern Scandinavia without admixture with local SHG-like populations, or local admixture of LBK-like farmers with a forager population that shows more similarity to WHG or Baltic hunter-gatherers19. A detailed joint analysis of genetic and archaeological data from hunter-gatherers from northern Germany, Denmark and the southern tip of Sweden is necessary to establish the role of local admixture during the emergence of the TRB culture. Our data support that the Neolithic PWC foragers are largely genetically continuous to SHG, which is congruent with their similarities in subsistence strategies, while continuity between EN TRB and PWC can also be seen in archaeological assemblages38 and can be attributed to contact between farmers and foragers. Indeed, genetic evidence of admixture between these groups shows that they were not completely isolated from each other but did likely not uphold continuous contact nor intermarry frequently during their prolonged parallel existence in Scandinavia. 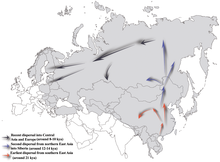 Estimated prehistoric migration routes for Y-chromosome haplogroup N lineage. In the archaeological understanding, the transition from Mesolithic to Neolithic in the Eastern Baltic region does not coincide with a large-scale population turnover and a stark shift in economy as seen in Central and Southern Europe. Rather, it is signified by a change in networks of contacts and the use of pottery, among other material, cultural and economic changes15. Our results suggest continued admixture between groups in the south of the Eastern Baltic region, who are more closely related to WHG, and northern or eastern groups, more closely related to EHG. Neolithic social networks from the Eastern Baltic to the River Volga could also explain similarities of the hunter-gatherer pottery styles, although morphologically analogous ceramics might also have developed independently due to similar functionality39. The genetic evidence for a change in networks and possibly even a large-scale population movement is most pronounced in the Middle Neolithic in individuals attributed to the CCC. The distribution of this culture overlaps in the north with the Narva culture and extends further north to Finland and Karelia. Its spread in the Eastern Baltic is linked with a significant change in imported raw materials, artefacts, and the appearance of village-like settlements15. We see a further population movement into the regions surrounding the Baltic Sea with the CWC in the Late Neolithic that was accompanied by the first evidence of extensive animal husbandry in the Eastern Baltic15. The presence of ancestry from the Pontic-Caspian Steppe among Baltic CWC individuals without the genetic component from north-western Anatolian Neolithic farmers must be due to a direct migration of steppe pastoralists that did not pick up this ancestry in Central Europe. It suggests import of the new economy by an incoming steppe-like population independent of the agricultural societies that were already established to the south and west of the Baltic Sea. The presence of direct contacts to the steppe could lend support to a linguistic model that sees an early branching of Balto-Slavic from a Proto-Indo-European language, for which the west Eurasian steppe was proposed as a homeland40,41,42. However, as farmer ancestry is found in later Eastern Baltic individuals, it is likely that considerable individual mobility and a network of contact throughout the range of the CWC facilitated its spread eastward, possibly through exogamous marriage practices43. Conversely, the appearance of mitochondrial haplogroup U4 in the Central European Late Neolithic after millennia of absence44 could indicate female gene-flow from the Eastern Baltic, where this haplogroup was present at high frequency.  Projected distributions of haplogroup N sub-haplogroups. (A) N*-M231, (B) N1*-LLY22g, (C) N1a-M128, (D) N1b-P43, (E) N1c-M46. Local foraging societies were, however, not completely replaced and contributed a substantial proportion to the ancestry of Eastern Baltic individuals of the latest LN and Bronze Age. This ‘resurgence’ of hunter-gatherer ancestry in the local population through admixture between foraging and farming groups recalls the same phenomenon observed in the European Middle Neolithic2,45 and is responsible for the unique genetic signature of modern-day Eastern Baltic populations. We suggest that the Siberian and East Asian related ancestry in Estonia, and Y-haplogroup N in north-eastern Europe, where it is widespread today, arrived there after the Bronze Age, ca. 500 calBCE, as we detect neither in our Bronze Age samples from Lithuania and Latvia. As Uralic speaking populations of the Volga-Ural region36 show high frequencies of haplogroup N36, a connection was proposed with the spread of Uralic language speakers from the east that contributed to the male gene pool of Eastern Baltic populations and left linguistic descendants in the Finno-Ugric languages Finnish and Estonian46,47. A potential future direction of research is the identification of the proximate population that contributed to the arrival of this eastern ancestry into Northern Europe. Nature Communicationsvolume 9, Article number: 442 (2018) |
|
|
|
Post by Admin on Oct 20, 2018 18:12:54 GMT
 Fig. 1. (A) Investigated skulls with strong (Left, AED_1108) intermediate, (Middle, STR_220), and no (Right, AED_92) skull deformation. (B) Location of archaeological sites from which genomes were analyzed (n = 41) on a map of Europe with today’s borders of Germany and the former borders of the western (green) and the eastern (light brown) Roman Empire. Bavarian sample sites (black square) are shown in the Inset. Sites in detail (number of deformed/nondeformed skulls): Altenerding (3/7), Alteglofsheim (1/0), Altheim (0/4), Barbing-Irlmauth (1/1), Burgweinting (1/1), and Straubing (3/9). Five additional individuals from Straubing were defined as intermediate. In addition, reference sites are given with which the Bavarian, Medieval genome data were compared: FN_2 (Freiham near Munich, ∼300 AD), deformed skull VIM_2 (Viminacium in Serbia, ∼550 AD), deformed skull KER_1 (Kerch on the Crimea, 256–401 cal AD), and two Sarmatians PR_4 and PR_10 (Pokrovka in southern Russia, 5th–2nd century BC). Europe experienced a profound cultural transformation between Late Antiquity and the Middle Ages that laid the foundations of the modern political, social, and religious landscape. During this period, colloquially known as the “Migration Period,” the Roman Empire gradually dissolved, with 5th and 6th century historiographers and contemporary witnesses describing the formation and migration of numerous Germanic peoples, such as the Goths, Alamanni, Gepids, and Longobards. However, the genetic and social composition of groups involved and the exact nature of these “migrations” are unclear and have been a subject of substantial historical and archaeological debate (1). In the mid 6th century AD, the historiographer Jordanes and the poet and hagiographer Venantius Fortunatus provide the first mention of a group known as the Baiuvarii that resided in modern day Bavaria. It is likely that this group had already started to form in the 5th century AD, and that it emanated from a combination of the romanized local population of the border province of the former Roman Empire and immigrants from north of the Danube (2). While the Baiuvarii are less well known than some other contemporary groups, an interesting archaeological feature in Bavaria from this period is the presence of skeletons with artificially deformed or elongated skulls (Fig. 1A). Artificial cranial deformation (ACD), which is only possible during early childhood, is a deliberate and permanent shaping of the head performed with great effort. In some societies reshaping the human skull has been seen as an ideal of beauty, while it may also have acted as a marker of status, nobility, or affiliation to a certain class or group (3). While ACD is a worldwide phenomenon that was practiced at least up to the 20th century (4), during the Late Roman and Early Medieval period in Europe it is most popularly associated with the Huns, an ambiguously defined nomadic group thought to have migrated into Europe from Asia (5). However, the earliest evidence for ACD appears in Europe in 2nd century AD burials in present day Romania that predate the proposed Hunnic invasion (6). Interestingly, there is a striking difference in the practice of ACD in the east and west of the continent: in eastern European burials, ACD is found at much higher frequency [50–80% of skulls in Hungary versus less than 5% in western graveyards] and is equally common among males and females and across all age classes (SI Appendix, Figs. S50–S52). In contrast, in Bavarian burials of the late 5th/early 6th century AD and elsewhere in western Europe, this cultural phenomenon is restricted to mainly adult females, while not noted in children and juveniles (SI Appendix, Fig. S53). Some scholars suggest these patterns are consistent with adult women migrating from eastern Europe or even Asia into western Europe (6). However, the associated assemblages of grave goods for women in the west with ACD cannot be clearly differentiated from non-ACD burials, leading others to argue that the practice was culturally adopted in western Europe from eastern foreign traditions (7). |
|
|
|
Post by Admin on Oct 21, 2018 18:19:33 GMT
 Fig. 2. Procrustes-transformed PCA of ancient samples using pseudohaploid calls based on off-target reads using an imputed POPRES modern reference dataset. Blue, green, and red male or female symbols are ancient Bavarian individuals with normal, intermediate, and elongated skulls, respectively. Orange circles are Anglo-Saxon era individuals. Large circles are medians for regions, dots are individuals. CE, central Europe; EE, eastern Europe; NE, northern Europe; NEE, northeastern Europe; NEW, northwestern Europe; SE, southern Europe; SEE, southeast Europe; WE, western Europe. Percentage of variation explained by PCs 1 and 2 for modern populations only is 0.25% and 0.15%. Geographical Origin and Genetic Diversity. A principal component analysis (PCA) of our ancient genomic data against that from a variety of modern reference population samples using both haploid and diploid datasets, as well as a supervised model-based clustering of the diploid data found that all men and 11 of the 13 women with normal skull shape clustered among modern northern and central European individuals (Figs. 2 and 3). Additional analyses examining patterns of haplotype sharing using only ancient whole genomes with coverage >10× also showed a high degree of matching to northern/central Europe relative to the alleged Roman soldier FN_2 (SI Appendix, Fig. S49), and as a group, both sexes with normal skull shape were closest (as assessed by FST on diploid 1-kb haplotypes) to modern 1000 Genomes samples of northern and western European Ancestry (CEU) (SI Appendix, Table S37).  Fig. 3. Supervised model-based clustering ADMIXTURE analysis for ancient samples based on phased haplotypes for individual 1,000 bp loci from the 5-Mb neutralome. Analysis is based on the best of 100 runs for K = 8, but NC_EUR is the ancestry summed across 1000 Genomes CEU, 1000 Genomes GBR, and GoNL populations (i.e., it represents a northern/central European ancestry). Blue, green, and red male or female symbols are ancient Bavarian individuals with normal, intermediate, and elongated skulls, respectively. A population assignment analysis (PAA) at the level of individual modern nation states suggested greatest genetic similarity of these normal-skulled individuals with modern Germans, consistent with their sampling location (Fig. 4 A and B and SI Appendix, Table S35). The only exceptions to this general pattern of northern/central European ancestry were the two women, STR_300 and STR_502, which were of a more southern ancestry associated with present day Greece and Turkey, respectively (SI Appendix, Fig. S29). |
|
|
|
Post by Admin on Oct 22, 2018 18:16:10 GMT
 Fig. 4. Geographic distribution of population assignment analysis (PAA) results on pseudohaploid calls from off-target reads summed across individuals for (A) all Bavarian males, (B) all Bavarian females with normal skulls, (C) all Bavarian females with elongated skulls, and (D) KER_1 and VIM_2. A much more diverse ancestry was observed among the females with elongated skulls, as demonstrated by a significantly greater group-based FIS (SI Appendix, Fig. S35). All these females had varying amounts of genetic ancestry found today predominantly in southern European countries [as seen by the varying amounts of ancestry inferred by model-based clustering that is representative of a sample from modern Tuscany, Italy (TSI), Fig. 3], and while the majority of samples were found to be closest to modern southeastern Europeans (Bulgaria and Romania, Fig. 4C), at least one individual, AED_1108, appeared to possess ∼20% East Asian ancestry (Fig. 3), which was also evident from the high number of haplotypes within the 5-Mb neutralome that were private to modern East Asian 1000 Genomes individuals (EAS), while also demonstrating an overall ancestry profile consistent with Central Asian populations (SI Appendix, Fig. S33). No modern European individual from the Simons Genome Diversity Panel (SGDP) (11) showed any evidence of significant East Asian ancestry except one Hungarian individual with less than 5%. A higher amount of East Asian ancestry was inferred for AED_1108 than all modern Caucasus and Middle Eastern individuals, and 28 of 33 South Asian individuals. From the individuals of intermediate skull size, one female (STR_310) exhibited the same southern European ancestry profile found in most other females with clear skull deformation, while the remaining four clustered clearly within northern/central Europeans (SI Appendix, Fig. S30). In addition, while females with normal skulls generally exhibited the same northern/central European component as the males (excluding STR_300 and STR_502), a small but significant East Asian component was consistently inferred for ALH_3. A diverse ancestry was also inferred for the two non-Bavarian samples with elongated heads. KER_1 from Ukraine possessed significant southern European ancestry as well as South Asian ancestry, with an overall profile that best matched modern Turkish individuals. The Gepid VIM_2 from Serbia demonstrated a similar Central Asian-like genetic profile to the Medieval Bavarian AED_1108 with an even larger East Asian component and number of private haplotypes but with less southern European/Middle Eastern ancestry (SI Appendix, Figs. S31 and S33). The two Sarmatian individuals (PR_4 and PR_10) fitted a general eastern European/western Asian profile, but also possessed a much larger northern European component [as represented by modern Finnish individuals (FIN)] similar to modern Russians, consistent with their sampling location. As previously observed in Schiffels et al. (12) contemporary Anglo-Saxon samples appeared to be primarily of northern/central European ancestry, with greatest similarity overall to modern British and Scandinavian individuals (SI Appendix, Fig. S32). A signal of population structure among our ancient samples was also observed when no modern reference samples were used to orientate genomic ancestry. An unsupervised model-based clustering analysis with K = 2 (SI Appendix, Fig. S27) essentially reiterated the results using the supervised analysis that identified individuals with predominantly northern/central versus southern European ancestry, while an outlier analysis identified STR_502, VIM_2, PR_10, KER_1, and AED_1108 as significantly distinct from all of the other samples, consistent with their significant non-European ancestry when orienting them with modern reference populations. It was also notable that no Bavarian individual (normal or ACD, male or female) possessed ancestry related to southwestern Europe, as represented by a sample of individuals sequenced from the Iberian population in Spain (IBS). This is in contrast to the Roman soldier dating to around 300 AD sampled from the same region, for which its largest ancestry component was IBS, with greatest genetic similarity to modern Spanish and southern French individuals (SI Appendix, Fig. S31). Based on an analysis of patterns of haplotype sharing, the Roman soldier (FN_2: 11.08×) was found to have substantially more southern European, West Asian, and Middle Eastern ancestry than two normal-skulled Early Medieval Bavarians with high genomic coverage (ALH_10: 12.17×, ALH_1: 13.27×) (SI Appendix, Figs. S48 and S49). |
|

