|
|
Post by Admin on Feb 23, 2022 21:05:29 GMT
Known as Dali, the mountainous site is located near the crossroads of cultural and genetic change during the Early and Late Bronze Age in a region that Frachetti calls the Inner Asia Mountain Corridor. At Dali, student researchers participating in the Washington University archaeological field school excavated skeletal samples that are a key part of the current ancient DNA study. The genetic results from burials at Dali show movement and admixture between groups with North Eurasian hunter gatherer ancestry and farmers from the Iranian plateau -- defining a bi-directional flow of ancestry along the mountains and steppes of Eurasia approximately 5000 years ago. 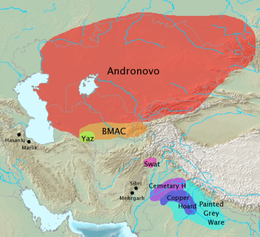 Separately, Frachetti and collaborators also sampled DNA from individuals of the Bactria-Margiana Archaeological Complex (or BMAC), a Bronze Age civilization of southern Central Asia, found in present-day Uzbekistan and Turkmenistan, which was discovered for the first time in the 1970s by Soviet archaeologists. Within the large number of samples from the BMAC included in the new genetic study (more than 100 individuals where previously there was no data), researchers were able to identify several key "outliers" -- individuals who reflect early steppe genetics, similar to those found at Dali -- living among an otherwise homogenous group. The discovery of a range of these genetically distinct individuals intermixed among regional populations fits with the archaeological evidence collected by Frachetti and his students over the last decade. They had previously determined that people in the large urban centers of the BMAC were trading goods with pastoral communities living along the mountains and nearby plains. "We had made the argument, as did many others, that steppe pastoralists were interacting with the BMAC on the basis of ceramics, metals, etc.," Frachetti said. "But there wasn't enough research, and the data were few. The new information we are getting from ancient DNA is critical to understanding the complex connectivity between these diverse communities across Asia." "What we see in both in the isotopic information as well as the archeological information is trade and exchange of agricultural material and production happening in both directions: north and south along this corridor," Harvard's Narasimhan said. "Now with the ancient DNA, we're actually seeing that in the people," he said. "It's giving us confidence to understand what happened in the past and how this process is happening."  Vivid narrative The new genetic results also open up a wide array of additional questions that archaeologists can now probe with new eyes. "The sort of questions you can ask now -- the sky's the limit," Frachetti said. "Working with this team has definitely opened up new ways of thinking for me. Questions about the degree to which trade and exchange versus important population movements brought innovation to various peoples, and how it shaped their future directions." "I've asked a lot of these questions for 25 years without the benefit of the genetics," Frachetti said. "So, it's exciting to return to my laundry list of questions and say, 'Wow, a lot of these are now made that much more crystal clear.'" The power of this study comes from its breadth and reach, made possible by samples from so many contributing archaeologists. "If you only have one archeologist help to tell the story, then likely that one might be inclined to support his or her own theories," Frachetti said. "But if, for example, you bring together data and ideas from 10, 20, 50 archaeologists -- many of whom have different, competing theories on the same topic -- it forces everybody to look at the science, and that's how progress is made! "That's really why teaming up with geneticists is so important. It's still not totally perfect in terms of how everyone wants things to be explained," Frachetti said. "But this paper brings us a rigorous, well-designed study with a massive baseline of ethically acquired data. Now all scholars can go back to it to focus on different sites or see how the data fit into their archaeological interpretation. "This study, and the important genetic record it establishes, allows us to shade in the narrative so much more vividly," he said. |
|
|
|
Post by Admin on Feb 24, 2022 2:10:42 GMT
The Formation of Human Populations in South and Central Asia Abstract By sequencing 523 ancient humans, we show that the primary source of ancestry in modern South Asians is a prehistoric genetic gradient between people related to early hunter-gatherers of Iran and southeast Asia. Following the Indus Valley Civilization’s decline, they mixed with people in the southeast to form one of the two main ancestral populations of South Asia whose direct descendants live in southern India. Simultaneously, they mixed with descendants of Steppe pastoralists who spread via Central Asia after 4000 years ago to form the other main ancestral population. The Steppe ancestry in South Asia has the same profile as that in Bronze Age Eastern Europe, tracking a movement of people that affected both regions and that likely spread the unique shared features shared between Indo-Iranian and Balto-Slavic languages.  The Bronze Age spread of Yamnaya steppe pastoralist ancestry into two subcontinents, Europe and South Asia. Pie charts reflect the proportion of Yamnaya ancestry, and dates reflect the earliest available ancient DNA with Yamnaya ancestry in each region. There is no ancient DNA yet for the ANI and ASI, so for these the range is inferred statistically. One Sentence Summary: Genome wide ancient DNA from 523 ancient individuals sheds light on genetic exchanges between the Steppe, Iran and South Asia, and highlights the parallel demographic histories of two subcontinents: Europe and South Asia. One Page Summary Introduction and Rationale: To elucidate the extent to which the major cultural transformations of farming, pastoralism and shifts in the distribution of languages in Eurasia were accompanied by movement of people, we report genome-wide ancient DNA data from 523 individuals spanning the last 8000 years mostly from Central Asia and northernmost South Asia. Results: Movements of people following the advent of farming resulted in genetic gradients across Eurasia that can be modeled as mixtures of seven deeply divergent populations. A key gradient formed in southwestern Asia beginning in the Neolithic and continuing into the Bronze Age, with more Anatolian farmer-related ancestry in the west and more Iranian farmer-related ancestry in the east. This cline extended to the desert oases of Central Asia and was the primary source of ancestry in peoples of the Bronze Age Bactria Margiana Archaeological Complex (BMAC). This supports the idea that the archaeologically documented dispersal of domesticates was accompanied by the spread of people from multiple centers of domestication. The main population of the BMAC carried no ancestry from Steppe pastoralists and did not contribute substantially to later South Asians. However, Steppe pastoralist ancestry appeared in outlier individuals at BMAC sites by the turn of the second millennium BCE around the same time as it appeared on the southern Steppe. Using data from ancient individuals from the Swat Valley of northernmost South Asia, we show that Steppe ancestry then integrated further south in the first half of the second millennium BCE, contributing up to 30% of the ancestry of modern groups in South Asia. The Steppe ancestry in South Asia has the same profile as that in Bronze Age Eastern Europe, tracking a movement of people that affected both regions and that likely spread the unique shared features shared between Indo-Iranian and Balto-Slavic languages. The primary ancestral population of modern South Asians is a mixture of people related to early Holocene populations of Iran and South Asia that we detect in outlier individuals from two sites in cultural contact with the Indus Valley Civilization (IVC), making it plausible that it was characteristic of the IVC. After the IVC’s decline, this population mixed with northwestern groups with Steppe ancestry to form the “Ancestral North Indians” (ANI) and with southeastern groups to form the “Ancestral South Indians” (ASI) whose direct descendants live today in tribal groups in southern India. Mixtures of these two post-IVC groups--the ANI and ASI--drive the main gradient of genetic variation in South Asia today. Conclusion: Earlier work recorded massive population movement from the Steppe into Europe early in the 3rd millennium BCE, likely spreading Indo-European languages. We reveal a parallel series of events leading to the spread of Steppe ancestry to South Asia, thereby documenting movements of people that were likely conduits for the spread of Indo-European languages. |
|
|
|
Post by Admin on Feb 25, 2022 3:18:17 GMT
Introduction The past ten thousand years have witnessed profound economic changes driven by the transition from foraging to food production, and have also witnessed dramatic changes in cultural practice evident from archaeology, the distribution of languages, and the written record. The extent to which these changes were associated with movements of people has been a mystery in Central Asia and South Asia in part because of a paucity of ancient DNA. We report genome-wide data from 523 individuals from Central Asia and northernmost South Asia from the Mesolithic period onward (1), and co-analyze them with previously published ancient DNA from across Eurasia and with data from diverse present-day people. In Central Asia, we studied the extent to which the spread of farming and herding practices from the Iranian plateau to the desert oases south of the Steppe was accompanied by movements of people or adoption of ideas from neighboring groups (2–4). For the urban communities of the Bactria Margiana Archaeological Complex (BMAC) in the Bronze Age, we assessed whether the people buried in its cemeteries descended directly from earlier smaller scale food producers, and also documented their genetic heterogeneity (5). Further to the north and east, we showed that the Early Bronze Age spreads of crops and domesticated animals between southwest Asia and eastern Eurasia along the Inner Asian Mountain Corridor (6) was accompanied by movements of people. Finally, we examined when descendants of the Yamnaya, who spread across the Eurasian Steppe beginning around 3300 BCE (7–9), began to appear in Central Asia south of the Steppe. In northernmost South Asia, we report a time transect of more than one hundred individuals beginning ~1200 BCE, which we co-analyze along with modern data from hundreds of present-day South Asian groups, as well as ancient DNA from neighboring regions (10). Previous analyses place the majority of present-day South Asians along a genetic cline (11) that can be modeled as having arisen from mixture of two highly divergent populations after 4000 years ago: the Ancestral North Indians (ANI) who harbor large proportions of ancestry related to West Eurasians, and the Ancestral South Indians (ASI) who are much less closely related to West Eurasians (12). We leveraged ancient DNA to place constraints on the genetic structure of the ANI and ASI and, in conjunction with other lines of evidence, to make inferences about when and where they formed. By modeling modern South Asians along with ancient individuals from sites in cultural contact with the IVC, we inferred a likely genetic signature for people of the Indus Valley Civilization (IVC) which reached its maturity in northwestern South Asia 2600–1900 CE. We also examined when Steppe pastoralist-derived ancestry (9) mixed with groups in South Asia, and placed constraints on whether Steppe-related ancestry or Iranian-related ancestry is more plausibly associated with the spread of Indo-European languages in South Asia. Dataset and Analysis Strategy We generated whole-genome ancient DNA data from 523 previously unsampled ancient individuals and increased the quality of data from 19 previously sequenced individuals. The individuals derive from three broad geographical regions: 182 from Iran and the southern part of Central Asia that we call Turan (present-day Turkmenistan, Uzbekistan, Tajikistan, Afghanistan and Kyrgyzstan), 209 from the Steppe and northern forest zone mostly in present-day Kazakhstan and Russia, and 132 from northern Pakistan. The ancient individuals are from 1) Mesolithic, Copper, Bronze and Iron Age Iran and Turan (12000–1 BCE from 19 sites) including the Bactria Margiana Archaeological Complex (BMAC); 2) early ceramic-using hunter-gatherers from the western Siberian forest zone who we show represent a point along an early Holocene cline of North Eurasians and who emerge as a valuable source population for modeling the ancestry of Central and South Asians (6400–3900 BCE from 2 sites); 3) Copper Age and Bronze Age pastoralists from the central Steppe, including from Bronze Age Kazakhstan (3400–800 BCE from 56 sites); and 4) northernmost South Asia, specifically Late Bronze Age, Iron Age and historical settlements in the Swat and Chitral districts of present-day Pakistan (~1200 BCE - 1700 CE from 12 sites) (Fig. 1, Table S1, (1, 13)). We prepared samples in dedicated clean rooms, extracted DNA (14, 15), and constructed libraries for Illumina sequencing (16, 17). We enriched the libraries for DNA overlapping around 1.2 million single nucleotide polymorphisms (SNPs) (7, 18, 19), sequenced the products on Illumina instruments, and performed quality control (Table S2) (7, 19, 20). Our final dataset after merging with previously reported data (7–9, 16, 18, 19, 21–31) spans 837 ancient individuals that passed all our analysis filters, which included removing individuals determined genetically to be first-degree relatives of other higher coverage individuals (Table S3), and restricting to the 92% of individuals (Table S1) that were represented by at least 15,000 of the targeted SNPs which we found was the number at which we began to be able to reliably estimate proportions of the deeply divergent ancestry sources. The median number of SNPs analyzed per individual was 617,000. We also merged with previously reported whole genome sequencing data from 686 present-day individuals (Table S1), and co-analyzed with 1,789 present-day people from 246 ethnographically-distinct groups in South Asia genotyped at ~600,000 SNPs (Table S5; (13)) (10, 32, 33).  Fig. 1 Overview of ancient DNA data. (A) Distribution of sites and associated archeological or radiocarbon dates along with the number of individuals meeting our analysis thresholds from each site. (B) Locations of ancient individuals for whom we generated ancient DNA that passed our analysis thresholds along with the locations of individuals from 140 groups from present-day South Asia that we analyzed as forming the Modern Indian Cline. Shapes distinguish the individuals from different sites. Data from 106 South Asian groups that do not fit along the Modern Indian Cline as well as AHG are not shown. (C) PCA analysis of ancient and modern individuals projected onto a basis formed by 1,340 present day Eurasians reflects clustering of individuals that mirrors their geographical relationships. An interactive version of this figure is presented in the Online Data Visualizer. We grouped individuals based on archaeological and chronological information, taking advantage of 269 direct radiocarbon dates generated on skeletal material from the individuals from whom we report DNA (Table S4). We further clustered individuals that were genetically indistinguishable within these groupings, and labeled outliers with ancestry that was significantly different from others at the same site and time period (13). For our primary analyses, we did not include individuals that were the sole representatives of their ancestry profiles, thereby reducing the chance that our conclusions were being driven by single individuals with contaminated DNA or misattributed archaeological context. This also ensured that each major analysis grouping was represented by many more SNPs that our minimum cutoff of 15,000 per individual. Thus, all but one analysis cluster included at least one individual covered by >200,000 SNPs, sufficient to support high resolution analysis of population history (19) (the exception is a pair of genetically similar outliers from the site of Gonur who are not the focus of any main analyses). We use Italic font to refer to genetic groupings and plain font to indicate archaeological cultures or sites. To make inferences about population structure, we began by carrying out principal component analysis (PCA) projecting ancient individuals onto the patterns of genetic variation in present-day Eurasians, a procedure that allowed us to obtain meaningful constraints on ancestry even of ancient individuals with limited coverage because each SNP from each individual can be compared to a large reference data set (34–36). This revealed three major clusters strongly correlating to the geographic regions of the Forest Zone/Steppe, Iran/Turan, and South Asia (Fig. 1), a pattern we replicate in ADMIXTURE unsupervised clustering (37). To test if groups of ancient individuals were heterogeneous in their ancestry, we used f4-statistics to measure whether different partitions of these groups into two subgroups differed in their degree of allele sharing to a third group (using a distantly related outgroup as a baseline). We also used f3-statistics to test for admixture (33). To model the ancestry of each group, we used qpAdm, which evaluates whether a tested group is consistent with deriving from a pre-specified number of source populations (relative to a set of outgroups), and if so estimates proportions of ancestry (7). We first used qpAdm to attempt to model groups from the Copper Age and afterward as a mixture of seven “distal” sources related deeply to pre-Copper Age or distantly related modern populations for which we have data (Box 1). In this paper we use the term ‘farmers’ to refer to people who either cultivated crops, or herded animals, or both; this definition covers not only large settled communities but also smaller and probably less sedentary communities like the early herders of the Zagros Mountains of western Iran from the site of Ganj Dareh. The latter kept domesticated animals but did not cultivate crops, and are a key reference population for this study as they had a distinctive ancestry profile that spread widely after the Neolithic (9, 24, 38). We also identified “proximal” models for each group as mixtures of temporally preceding groups (10). We implemented an algorithm, DATES, for estimating the age of population mixtures by measuring the average size of segments of ancestry derive from the admixing populations, an approach whose reliability we verified by computer simulation (10) and that is an improvement relative to methods not optimized for analysis of ancient DNA (33, 39) (the approach’s robustness reflects the fact that it relies for its molecular clock on the accurately measured rate of meiotic recombination in humans (40)). In Box 2, we summarize the findings of these analyses (we use the same headings in Box 2 and the main text to allow cross-referencing), while the Online Data Visualizer (1) allows an interactive exploration of the data. |
|
|
|
Post by Admin on Feb 25, 2022 21:19:47 GMT
Box 1.
Seven Source Populations Used for Distal Ancestry Modeling
Anatolia_N Anatolian farmer-related: Represented by 7th millennium BCE western Anatolian farmers (19)
Ganj_Dareh_N Iranian early farmer-related: Represented by 8th millennium BCE early goat herders from the Zagros Mountains of Iran (9, 24)
WEHG Western European Hunter-Gatherer-related: represented by 9th millennium BCE Western Europeans (7, 19, 32, 69). (WEHG and EEHG discussed below were denoted WHG and EHG in previous studies, but as we co-analyze them with hunter-gatherers from Asia we modify the names to specify a European origin.)
EEHG Eastern European Hunter-Gatherer-related: represented by 6th millennium BCE hunter-gatherers from Eastern Europe (19, 32)
WSHG West Siberian Hunter-Gatherer-related: a previously undescribed deep source of Eurasian ancestry represented in this study by three individuals from the Forest Zone of Central Russia dated to the 6th millennium BCE.
ESHG East Siberian Hunter-Gather-related: represented by 6th millennium BCE hunter-gatherers from the Lake Baikal region with ancestry deeply related to East Asians (26)
AHG Andamanese Hunter-Gatherer-related: represented by present day indigenous Andaman Islanders (55) who we hypothesize are related to unsampled indigenous South Asians (Ancient Ancestral South Indians - AASI)
Box 2.
Summary of Key Findings
Iran and Turan
A West-to-East Cline of Decreasing Anatolian Farmer-Related Ancestry. There was a west-to-east gradient of ancestry across Eurasia in the Copper Age and Bronze Ages—the Southwest Asian Cline—with more Anatolian farmer-related ancestry in the west and more WSHG- or AASI-related ancestry in the east, superimposed on primary ancestry related to early Iranian farmers. The establishment of this gradient correlates in time to the spread of plant-based agriculture across this region, raising the possibility that people of Anatolian ancestry spread this technology east just as they helped spread it west into Europe.
People of the BMAC Were Not a Major Source of Ancestry for South Asians. The primary BMAC population largely derived from preceding local Copper Age peoples who were in turn closely related to people from the Iranian plateau, and had little of the Steppe ancestry that is ubiquitous in South Asia today.
Steppe Pastoralist-Derived Ancestry Arrived in Turan by 2100 BCE. We find no evidence of Steppe pastoralist-derived ancestry in groups at BMAC sites prior to 2100 BCE, but multiple outlier individuals buried at these sites show that by ~2100–1700 BCE, BMAC communities were regularly interacted with peoples carrying such ancestry.
An Ancestry Profile Widespread During the Indus Valley Civilization. We document a distinctive ancestry profile—45–82% Iranian farmer-related and 11–50% AASI (with negligible Anatolian farmer-related admixture)—that was present at two sites in cultural contact with the Indus Valley Culture (IVC). Combined with our detection of this same ancestry profile (in mixed form) about a millennium later in the post-IVC Swat Valley, this documents an Indus Periphery Cline during the flourishing of the IVC. Ancestors of this group formed by admixture ~5400–3700 BCE. There is little if any Anatolian farmer-related ancestry in the Indus Periphery Cline.
The Steppe and Forest Zone
Ancestry Clines in North Eurasia Established After the Advent of Farming. Prior to the spread of farmers and herders, northern Eurasia was characterized by a west-to-east gradient of very divergent hunter-gatherer populations with increasing proportions of relatedness to present-day East Asians: from Western European Hunter-Gatherers (WEHG), to Eastern European Hunter-Gatherers (EEHG), to West Siberian Hunter-Gatherers (WSHG), to Eastern Siberian Hunter-Gatherers (ESHG). Mixture of people along this ancestry gradient and its counterpart to the south formed five later clines following the advent of farming, the three northern ones of which are the European Cline, the Caucasus Cline, and the Central Asian Cline.
A Distinctive Ancestry Profile Stretching from Eastern Europe to Kazakhstan in the Bronze Age. We add more than one hundred samples from the previously described Western_Steppe_MLBA genetic cluster, including individuals associated with the Corded Ware, Srubnaya, Petrovka, and Sintashta archaeological complexes, and characterized by a mixture of about two-thirds ancestry related to Yamnaya Steppe pastoralists (from the Caucasus Cline), and European farmers (from the European Cline) suggesting that this population formed at the geographic interface of these two groups in Eastern Europe. Our analysis suggests that in the central Steppe and Minusinsk Basin in the Middle to Late Bronze Age, Western_Steppe_MLBA ancestry mixed with about 9% ancestry from previously established people from the region carrying WSHG-related to form a distinctive Central_Steppe_MLBA cluster that was the primary conduit for spreading Yamnaya Steppe pastoralist-derived ancestry to South Asia.
Bidirectional Mobility Along the Inner Asian Mountain Corridor. Beginning in the 3rd millennium BCE and intensifying in the 2nd millennium BCE, we observe multiple individuals in the Central Steppe who lived along the Inner Asian Mountain Corridor and who harbored admixture from Turan, documenting northward movement into the Steppe in this period. By the end of the 2nd millennium BCE, these people were later joined by numerous outlier individuals with East Asian-related admixture which become ubiquitous in the region by the Iron Age (25, 52). This ancestry is also seen in later groups with known cultural impacts on South Asia including Huns, Kushans and Sakas and is hardly present in the two primary ancestral populations of South Asia, suggesting that the Steppe ancestry widespread in South Asia derived from pre-Iron Age Central Asians.
South Asia
Three Ancestry Clines That Succeeded Each Other in Time in South Asia. We identify a unique trio of source populations that fits geographically and temporally diverse South Asians since the Bronze Age: a mixture of AASI, an Indus Periphery Cline group with predominantly Iranian farmer-related ancestry, and Central_Steppe_MLBA. Two-way clines driven more by populations that were mixtures of these three sources succeeded each other in time: prior to 2000 BCE the Indus Periphery Cline had no detectable Steppe ancestry, beginning after 2000 BCE the Steppe Cline, and finally the Modern Indian Cline.
The ASI and ANI Arose as Indus Periphery Cline People Mixed with Groups to the North and East. An ancestry gradient of which the Indus Periphery Cline individuals were a part played a pivotal role in the formation of both the two proximal sources of ancestry in South Asia: a minimum of ~55% Indus Periphery Cline ancestry for the ASI and ~70% for the ANI. Today there are groups in South Asia with very similar ancestry to the statistically reconstructed ASI suggesting that they have essentially direct descendants today. Much of the formation of both the ASI and ANI occurred in the 2nd millennium BCE. Thus, the events that formed both the ASI and ANI overlapped the time of the decline of the IVC.
Steppe Ancestry in South Asia is Primarily from Males and Disproportionately High in Brahmins. Most of the Steppe ancestry in South Asia derives from males, pointing to asymmetric social interaction between descendants of Steppe pastoralists and peoples of the Indus Periphery Cline. Groups that view themselves as being of traditionally priestly status, including traditional custodians of liturgical texts in the early Indo-European language Sanskrit, tend (with exceptions) to have more Steppe ancestry than expected based on ANI-ASI mixture, providing an independent line of evidence for a Steppe origin for South Asia’s Indo-European languages.
|
|
|
|
Post by Admin on Feb 26, 2022 3:23:55 GMT
Iran and Turan A West-to-East Cline of Decreasing Anatolian Farmer-Related Ancestry We studied the genetic transformations accompanying the spread of agriculture eastward from Iran beginning in the 7th millennium BCE (41, 42). We replicate previous findings that 9th to 8th millennium BCE herders from the Zagros Mountains of western Iran harbored a distinctive West Eurasian-related ancestry profile (9, 27), while later groups across a broad region were admixed between this ancestry and that related to early Anatolian farmers. Our analysis reveals a west-to-east cline of decreasing Anatolian farmer-related admixture in the Copper Age and Bronze Age ranging from ~70% in Anatolia to ~31% in eastern Iran, to ~7% in far eastern Turan (Fig. 1; (13), Fig. S10, Table S8–S16). This suggests that the archaeologically documented spread of a shared package of plants and animal domesticates from diverse locations across this region was accompanied by bi-directional spread of people and mixture with the local groups they encountered (3, 41, 43, 44). We call this the Southwest Asian Cline. In the far east of the Southwest Asian Cline (eastern Iran and Turan) in individuals from the 3rd millennium BCE, we detect not only the smallest proportions of Anatolian farmer-related admixture but also admixture related to West Siberian Hunter Gatherers (WSHG) (plausibly reflecting admixture from unsampled hunter-gatherer groups who inhabited this region prior to the spread of Iranian farmer-related ancestry into it). This shows that North Eurasian-related ancestry impacted Turan well before the spread of descendants of Yamnaya Steppe pastoralists into the region. We can exclude the possibility that the Yamnaya were the source of this North Eurasian-related ancestry, as they had more EEHG-than WSHG-related ancestry, and also carried high frequencies of mitochondrial DNA haplogroup type U5a as well as Y chromosome haplogroup types R1b or R1a not represented in Iran and Turan in this period ((13), Table S93-S94). People of the BMAC Were Not a Major Source of Ancestry for South Asians From Bronze Age Iran and Turan, we obtained genome-wide data for 84 ancient individuals (3000–1400 BCE) who lived in four urban sites of the Bactria Margiana Archaeological Complex (BMAC) and its immediate successors. The great majority of these individuals fall in a cluster genetically similar to the preceding groups in Turan, consistent with the hypothesis that the BMAC coalesced from preceding pre-urban populations (5). We infer three primary genetic sources: early Iranian farmer-related ancestry (~60–65%), and smaller proportions of Anatolian farmer- (~20–25%) and WSHG-related ancestry (~10%). Unlike preceding Copper Age individuals from Turan, people of the BMAC cluster also harbored an additional 2–5% ancestry related (deeply in time) to Andamanese Hunter-Gatherers (AHG). This evidence of north-to-south gene flow from South Asia is consistent with the archaeological evidence of cultural contacts between the Indus Valley Civilization and the BMAC and the existence of an IVC trading colony in northern Afghanistan (although we lack ancient DNA from that site) (45), and stands in contrast to our qpAdm analyses showing that a reciprocal north-to-south spread is undetectable. Specifically, our analyses reject the BMAC and the people who lived before them in Turan as plausible major sources of ancestry for diverse ancient and modern South Asians by showing that their ratio of Anatolian farmer-related to Iranian farmer-related ancestry is too high for them to be a plausible source for South Asians (p<0.0001, χ2 test; (13), Fig S50–S51). A previous study (26) fit a model in which a population from Copper Age Turan was used a source of the Iranian farmer-related ancestry in present-day South Asians, thus raising the possibility that the people of the BMAC whom the authors correctly hypothesized were primarily derived from the groups that preceded them in Turan were a major source population for South Asians. However, that study only had access to 2 samples from this period compared to the 36 we report with this study, and it lacked ancient DNA from individuals from the BMAC period or from any ancient South Asians. With additional samples, we have the resolution to show that none of the large number of Bronze and Copper Age populations from Turan for which we have ancient DNA fit as a source for the Iranian farmer-related ancestry in South Asia. Steppe Pastoralist-Derived Ancestry Arrived in Turan by 2100 BCE Our large sample sizes from Central Asia, including individuals from BMAC sites, are a particular strength of this study, allowing us to detect outlier individuals with ancestry different from those living at the same time and place, and revealing cultural contacts that would be otherwise difficult to appreciate (Fig. 2). Around ~2300 BCE, we observe three outliers in BMAC-associated sites carrying WSHG-related ancestry and we report data from the third millennium BCE from three sites in Kazakhstan and one in Kyrgyzstan that fit as sources for them (related ancestry has been found in ~3500 BCE Botai culture individuals (26)). Yamnaya-derived ancestry arrived by 2100 BCE, since from 2100–1700 BCE we observe outliers from three BMAC-associated sites carrying ancestry ultimately derived from Western_Steppe_EMBA pastoralists, in the distinctive admixed form typically carried by many Middle to Late Bronze Age Steppe groups (with roughly two thirds of the ancestry being of Western_Steppe_EMBA origin, and the rest consistent with deriving from European farmers). Thus, our data document a southward movement of ancestry ultimately descended from Yamnaya Steppe pastoralists that spread into Central Asia by the turn of the 2nd millennium BCE.  Fig. 2. Outlier analysis reveals ancient contacts between sites. We plot the average of Principal Component 1 (x-axis) and Principal Component 2 (y-axis) for the West Eurasian and All Eurasian PCA plots, as we found that this aids visual separation of the ancestry profiles. (A) In the Middle to Late Bronze Age Steppe, we observe in addition to the Western_Steppe_MLBA and Central_Steppe_MLBA clusters (indistinguishable in this projection), outliers admixed with other ancestries. The BMAC-related admixture in Kazakhstan documents northward gene flow onto the Steppe and confirms the Inner Asian Mountain Corridor as a conduit for movement of people. (B) At Shahr-i-Sokhta in eastern Iran, there are two primary groupings: one with ~20% Anatolian farmer-related ancestry and no detectable AHG-related ancestry, and the other with ~0% Anatolian farmer-related ancestry and substantial AHG-related ancestry (Indus Periphery Cline). (C) In individuals of the BMAC and successor sites, we observe a main cluster as well as numerous outliers: outliers >2000 BCE with admixture related to WSHG, outliers >2000 BCE on the Indus Periphery Cline (with an ancestral similar similar to the outliers at Shahr-i-Sokhta), and outliers after 2000 BCE that reveal how Central_Steppe_MLBA ancestry had arrived. (D) In the Late Bronze Age and Iron Age of northernmost South Asia, we observe a main cluster consistent with admixture between peoples of the Indus Periphery Cline and Central_Steppe_MLBA, and variable Steppe pastoralist-related admixture. |
|

