|
|
Post by Admin on Mar 3, 2023 3:36:51 GMT
Before the LGM The Gravettian culture was one of the most widely distributed Upper Palaeolithic cultures across western Eurasia before the LGM19. It is often considered as a pan-European cultural mosaic with regional variations in material to symbolic productions20,21. In this debated framework, Gravettian-associated individuals have been suggested to represent a biologically homogeneous population on the basis of craniometric and genomic data4,22. However, published Gravettian-associated genomes originate from central and southern Europe, leaving the genetic profile of Gravettian-associated human groups from western and southwestern Europe undescribed. To gain an overview of the genomic background of European hunter-gatherers before the LGM, we used multidimensional scaling (MDS) to plot a dissimilarity matrix of pairwise outgroup f3-statistics in the form 1 − f3(Mbuti; pop1, pop2) (Fig. 2a). This plot reveals the presence of three distinct groupings: (1) a pre-40 ka group with individuals from the Ust’Ishim, Bacho Kiro, Zlatý kůň and Peştera cu Oase sites, (2) a Věstonice cluster including Gravettian-associated individuals from central–eastern and southern European sites (Dolní Věstonice, Pavlov, Krems-Wachtberg, Paglicci and Ostuni), and (3) a Fournol cluster (hereafter, Fournol cluster or ancestry) comprising Gravettian-associated individuals from western and southwestern European sites (Ormesson, La Rochette, Fournol and two Serinyà cave sites (Mollet III and Reclau Viver)). The previously described Věstonice cluster, including a newly reported 29,000-year-old individual from Paglicci cave (Paglicci 12) in southern Italy, is closely related to the previously published genomes from Sunghir and Kostenki 12 in western Russia, which are dated to 34 ka and 32 ka, respectively4,23. The newly defined Fournol cluster is closely related to Aurignacian-associated individuals from Belgium dated to 35 ka (Goyet Q116-1 and the newly reported Goyet Q376-3 individual). Notably, and contrary to the report by Fu et al.4, another Gravettian-associated population from central–western Europe (Goyet in Belgium, n = 6 individuals) is both geographically and genetically intermediate between the Věstonice and Fournol clusters. The similarity between Goyet Q116-1 and Goyet Q376-3 and the Fournol cluster is also observed at the mtDNA level, with both groups including individuals who carried mtDNA haplogroup M, which has not been found in European individuals from after the LGM24 (Extended Data Figs. 1 and 2). Fig. 2: Genetic differences among Gravettian-associated populations. 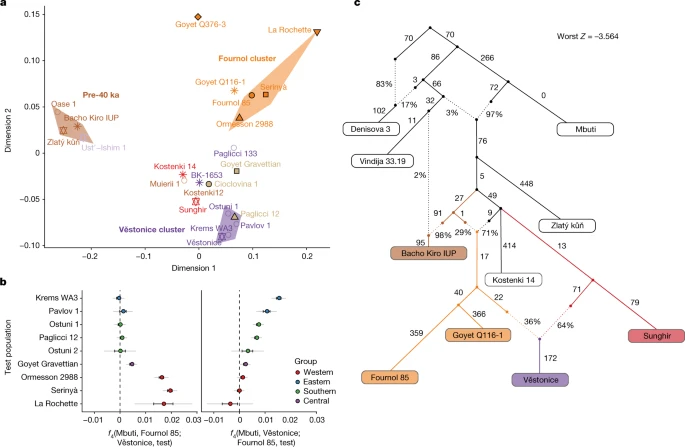 a, MDS plot of pre-LGM individuals. The pre-40 ka group and the Fournol and Věstonice clusters are marked as shaded areas in different colours. Individuals and groups are plotted with the same colours and symbols as in Fig. 1 and names are indicated next to the symbols. b, Gravettian-associated individuals form two distinct groups, with central-eastern and southern European individuals as part of the Věstonice cluster and western and southwestern European individuals as part of the Fournol cluster. In central-western Europe, Gravettian-associated individuals from Goyet show affinity to both clusters. Error bars show 1× s.e.m. (black) or 3× s.e.m. (grey) of the f4 values estimated from 5 cM-block jackknife analysis. c, Admixture graph modelling of the main pre-LGM European hunter-gatherer lineages created using qpGraph. We further validated the genetic distinction between the Věstonice and Fournol clusters observed in the MDS plot with a series of f4-statistics (Supplementary Data 2.B). All individuals belonging to the Fournol cluster show higher affinity to Goyet Q116-1 than to the Sunghir group (n = 4), and the Věstonice-cluster individuals show higher affinity to the Sunghir group than to Goyet Q116-1 (Extended Data Fig. 3). These f4-statistics also confirm that Goyet Q376-3 carries a similar ancestry to Goyet Q116-1 and Kostenki 12 carries a similar ancestry to the Sunghir group, whereas Bacho Kiro 1653 (35 ka) from Bulgaria, Muierii 1 (34 ka) and Cioclovina 1 (32 ka) from Romania, and Paglicci 133 (33 ka) from southern Italy are equally related to Goyet Q116-1 and Sunghir. We further tested whether individuals included in the Věstonice and Fournol clusters share similar allele frequencies with the main representatives of those two clusters. With the statistics f4(Mbuti, Fournol 85; Věstonice, test) and f4(Mbuti, Věstonice; Fournol 85, test), we show that all Věstonice-cluster individuals are significantly closer (|Z|>3) to the Věstonice group (n = 5) and the Fournol-cluster individuals are closer to Fournol 85, whereas the geographically intermediate Gravettian-associated Goyet group shows extra affinity to both clusters (Fig. 2b). We further modelled the genetic profile of pre-LGM individuals with qpGraph (Supplementary Information, section 10 and Supplementary Figs. 19–25). The admixture graph shows that the Bacho Kiro IUP group (n = 3) shares ancestry with multiple early modern human lineages2 (Supplementary Information, section 7), and that the more than 45,000-year-old Zlatý kůň genome1 is the most deeply divergent non-African lineage sequenced to date (Extended Data Fig. 4). This is also validated by f4-statistics of the form f4(Mbuti, Zlatý kůň; test1, test2), which are consistent with zero for all other pre-LGM hunter-gatherers (Supplementary Data 2.C), indicating an equidistant relationship of Zlatý kůň to the tested groups. When Gravettian-associated individuals are included in an admixture graph also featuring Kostenki 14, we find that Fournol 85 fits best as a sister lineage of Goyet Q116-1, whereas the Věstonice group is modelled as a mixture between a lineage related to the Sunghir group and one related to the Goyet Q116-1–Fournol 85 branch (Fig. 2c). This is also supported by f4-statistics of the form f4(Mbuti, Fournol 85; Sunghir, test), which are significantly positive for all the individuals included in the Věstonice cluster (Supplementary Data 2.B). Therefore, as previously reported2, the Věstonice cluster itself results from admixture between western and eastern lineages, which might contribute to the observed homogeneity in cranial morphology among Gravettian-associated individuals22. These results show that some, but not all, of the genomic ancestries present in Europe between around 40 ka and 30 ka survived in the Gravettian-associated populations studied so far. The Kostenki (and Sunghir group) ancestry contributed to the previously described Věstonice cluster represented by Gravettian-associated individuals from central-eastern and southern Europe4. By contrast, the Goyet Q116-1 genetic profile gave rise to the newly described Fournol cluster identified in Gravettian-associated individuals from western and southwestern Europe. Notably, this genetic distinction coincides with dissimilarities in mortuary practice among genetically analysed Gravettian-associated individuals from different parts of Europe. Individuals in western and southwestern Europe related to the Fournol cluster are consistently deposited in cave sites and occasionally exhibit anthropogenic marks whereas individuals related to the Věstonice cluster are buried with grave goods and/or personal ornaments and ochre in open air or cave sites in central-eastern and southern Europe, respectively (Supplementary Figs. 29–32 and Supplementary Table 4). The oldest individual in the Fournol cluster is Ormesson 2988 from northeastern France (31 ka, Early/Middle Gravettian), whereas a Gravettian group from Goyet in Belgium (27 ka, Late Gravettian) is found to be a mixture between the Věstonice and Fournol clusters. This suggests that between the Early/Middle and Late Gravettian there was an east-to-west expansion of the Věstonice-associated ancestry that reached central-western Europe and created a longitudinal admixture cline between those two genetically distinct pre-LGM populations. |
|
|
|
Post by Admin on Mar 8, 2023 19:20:23 GMT
LGM in southwestern and western Europe The Solutrean culture is temporally intermediate between the Gravettian and the Magdalenian (or the Badegoulian) cultures, and is found in southwestern and western Europe, which are considered to have been climatic refugia for human populations during the LGM25,26. However, the extent to which groups associated with the Solutrean culture are in genetic continuity with earlier and later populations from the same region is unknown because no genomic data from Solutrean-associated individuals have been reported previously. Both newly sequenced genomes from Solutrean-associated individuals (Le Piage II (23 ka) from southwestern France and La Riera (level 14, 21 ka) from northern Spain) show a generalized affinity with members of the Fournol and GoyetQ2 clusters in outgroup f3-statistics (Supplementary Data 2.A). In the MDS plot, the Le Piage II individual falls particularly close to individuals belonging to the Fournol cluster, suggesting a local genetic continuity of this ancestry during the LGM (Supplementary Fig. 13). F4-statistics further support this view, revealing that Le Piage II is more closely related to the Fournol cluster than the Věstonice cluster (f4(Mbuti, Le Piage II; Věstonice, Fournol 85) ≫ 0, Z = 6.58). We also compared its affinity to El Mirón (northern Spain), the oldest Magdalenian-associated individual sequenced to date (19 ka). F-statistics suggest that Le Piage II is genetically intermediate between Fournol 85 and El Mirón (Supplementary Data 2.D). Moreover, previous studies have shown that El Mirón carries a genetic contribution from the Villabruna cluster, which is found in Epigravettian-associated individuals from Italy4,15. El Mirón has a significantly higher similarity to the Villabruna cluster than Fournol 85 and Le Piage II, while the affinity to the Villabruna cluster in Le Piage II is not significantly higher than in Fournol 85 (Supplementary Data 2.D). Overall, the Solutrean-associated Le Piage II individual links the preceding Fournol ancestry with the succeeding ancestry found in El Mirón, providing direct evidence for genetic continuity throughout the LGM in southwestern and western Europe. These European regions, therefore, constitute climatic refugia where human populations survived during the LGM. Post-LGM in the Italian peninsula After the LGM, the Epigravettian culture was widespread in southern and southeastern Europe. In spite of growing discussions about its nature27,28, the Epigravettian culture has been traditionally assumed to be the result of a transition from the preceding local Gravettian29. However, the level of genetic continuity between individuals associated with these cultures and the population structure among Epigravettian-associated individuals have not been fully explored. Here, we report genomic data from 4 individuals, including 3 approximately 13,000-year-old genomes from northeastern Italy (Pradis 1), northwestern Italy (Arene Candide 16) and Sicily (San Teodoro 2), as well as increased genome-wide coverage from Tagliente 215 dated to 17 ka. In the MDS plot, we find that all of the newly and previously reported Epigravettian-associated individuals fall within the Villabruna cluster4 (Fig. 1c). A series of f4-symmetry statistics confirm that all the Epigravettian-associated individuals are cladal, and do not share excess affinity with any local (Paglicci 12) or non-local preceding ancestries (Goyet Q116-1, Kostenki 14, Mal’ta 1 or Věstonice) (Supplementary Data 2.F). Moreover, none of the Epigravettian-associated individuals have more affinity to southern European than to central-eastern European Gravettian-associated groups, as shown by f4(Mbuti, Epigravettian-associated individual/group; Věstonice, Paglicci 12) that is consistent with 0 (Supplementary Data 2.G). Next, we investigated the genetic relationships between Epigravettian-associated individuals across the Italian peninsula, by reconstructing a phylogeny based on a matrix of pairwise f2 genetic distances (Fig. 3a and Supplementary Fig. 9) and testing the relative affinity among them using f4-statistics in the form f4(Mbuti, Epigravettian A; Epigravettian B, Epigravettian C) (Supplementary Data 2.E). The inferred topology reveals a phylogeographic pattern irrespective of individual ages. In particular, the 13 ka Pradis 1 individual from northeastern Italy represents the most basal lineage compared to all other Epigravettian-associated individuals, including the older Tagliente 2 and Villabruna genomes from northern Italy (group 1). Individuals from northwestern Italy (Arene Candide 16), central Italy (Continenza) and Sicily fall on a phylogenetically more derived branch (group 2), which further diversified into a branch composed of Sicilian hunter-gatherers only (group 3). Within Sicily, the 14 ka Oriente C individual shows higher affinity with the much younger but geographically closer 10 ka Uzzo group30 (n = 2) than with the almost contemporaneous San Teodoro 2 individual from eastern Sicily. Fig. 3: The population substructure and diversity of Epigravettian-associated groups in southern Europe. 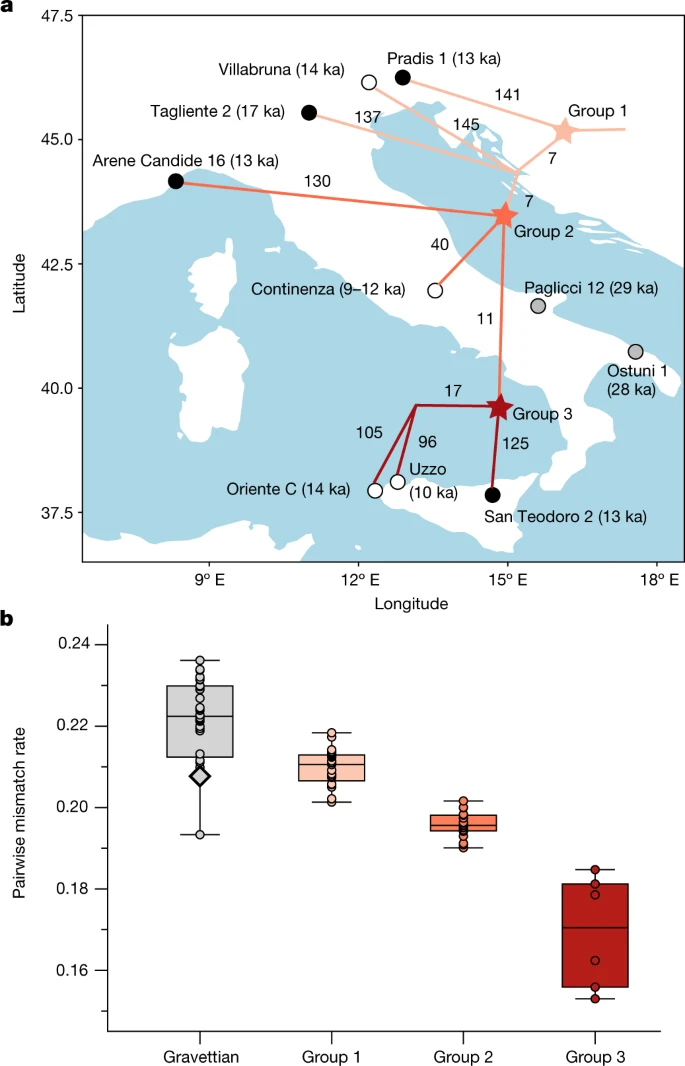 Finally, we estimated the genetic diversity of Epigravettian-associated individuals in the dataset by calculating both pairwise mismatch rates (PMR) on pseudo-haploid genotypes and individual heterozygosity levels on pseudo-diploid genotypes (Supplementary Data 3.A). Compared with the genetic diversity observed among all analysed Gravettian-associated groups, Epigravettian-associated individuals show significantly lower amounts of genetic diversity (two-tailed t-test, P < 0.001) (Fig. 3b). Moreover, we reveal a north-to-south decrease in genetic diversity among the Epigravettian-associated groups, with the highest PMR and heterozygosity values found in northern Italian individuals (group 1), intermediate in western and central Italian individuals (group 2) and the lowest in Sicilian individuals (group 3) (Fig. 3b). A similar pattern is observed through the analysis of ROH segments (Extended Data Fig. 5 and Supplementary Information, section 9). We detect the highest amount of ROHs in Epigravettian-associated individuals from Sicily, who carry an extreme amount of more than 200 cM of short ROHs (4–8 cM). This suggests a very small recent effective population size, estimated to be in the order of around 70 individuals (Supplementary Table 2), causing the low genetic diversity in Sicilian Epigravettian hunter-gatherers. To summarize, our results highlight a genetic turnover in the Italian peninsula of the Gravettian-associated Věstonice cluster by the Epigravettian-associated Villabruna cluster that might correlate with discontinuities observed in the archaeological record31. We show that all analysed Epigravettian-associated individuals carry a homogeneous Villabruna ancestry, with the intra-group genetic structure mainly determined by their geographical, and not temporal, distribution. The phylogenetic reconstruction of Epigravettian-associated genomes, with Pradis 1 diverging more deeply than all others, indicates that the turnover took place much earlier than 17 ka—the date of the more derived Tagliente 2 genome. This, together with the evidence of Villabruna ancestry in El Mirón 19 ka, further suggests that this genetic discontinuity could be the result of palaeogeographic and palaeoecological transformations connected to the LGM32, rather than to the Bølling–Allerød warming period4,15 (14.7–12.9 ka). In addition, our phylogeographic analysis points to northeastern Italy as the possible entry point of the Epigravettian-associated gene pool in the Italian peninsula. This finding, in conjunction with the genetic affinity of the Villabruna cluster to ancient and present-day Near Eastern ancestries4,15,16 (Supplementary Information, section 8, Supplementary Fig. 18 and Supplementary Data 2.O), suggests the Balkans as a source of the incoming Epigravettian-associated population. The LGM could thus have created a corridor south of the Alps for east-to-west human movements that genetically connected hunter-gatherer populations from the Balkans to Iberia, possibly also via dispersals along existing lower-sea-level coasts32. |
|
|
|
Post by Admin on Mar 9, 2023 18:25:25 GMT
Post-LGM in western and central Europe The Magdalenian culture was widely distributed in southwestern, western and central Europe after the LGM33. Despite this wide geographical range, it is not clear whether different groups associated with this culture originated from a common source population and how those groups were genetically related to each other. Previous studies identified two different genetic compositions in Magdalenian-associated individuals—the GoyetQ2 cluster including central-western European genomes dated to around 15 ka (from France, Belgium and Germany), and the ancestry of the El Mirón individual from Spain4,14 from around 19 ka. Both of these ancestries carry a genetic component distantly related to the Goyet Q116-1 individual dated to 35 ka, with the Iberian individual also showing an affinity to the Villabruna cluster4,14. By co-analysing previously published data with our newly reported genomes associated with the Magdalenian from La Marche (18 ka) and Pincevent (15 ka) in western and northern France, respectively, and Maszycka (18–16 ka) in southern Poland, we confirm that the Goyet Q116-1 ancestry survived in all studied Magdalenian-associated genomes besides in Gravettian and Solutrean-associated individuals from southwestern and western Europe (Fig. 1). Notably, the Fournol ancestry provides a better proxy than Goyet Q116-1 for the genetic component found in the GoyetQ2 cluster and in El Mirón (Supplementary Data 2.H). However, using f4-statistics, we show that all Magdalenian-associated individuals, and not only El Mirón, carry Villabruna-related ancestry when compared to the Fournol cluster (Supplementary Data 2.H). This affinity is even stronger towards Epigravettian-associated individuals from western and central Italy and Sicily (group 2 and group 3, respectively) than to those from northern Italy (group 1) (Supplementary Data 2.F). We thus modelled individuals belonging to the GoyetQ2 cluster and El Mirón as a mixture between the Fournol 85 and Arene Candide 16 genomes as proxies to represent the Fournol and Villabruna ancestries, respectively, in Magdalenian-associated groups (Fig. 4a). Besides El Mirón, who has around 43% Villabruna ancestry, all other Magdalenian-associated individuals have a lower proportion of this component (19–29%) and can thus be assigned to the GoyetQ2 cluster (Fig. 4a and Supplementary Data 3.C). This is further validated by f4-statistics of the form f4(Mbuti, Arene Candide 16; Goyet Q-2, Magdalenian-associated individuals), which is significantly positive only for El Mirón, whereas all other tested individuals and Goyet Q-2 are symmetrically related with respect to Arene Candide 16 (Supplementary Fig. 26 and Supplementary Data 2.H). Fig. 4: Ancestry modelling of post-19 ka individuals in southwestern, western and central Europe. 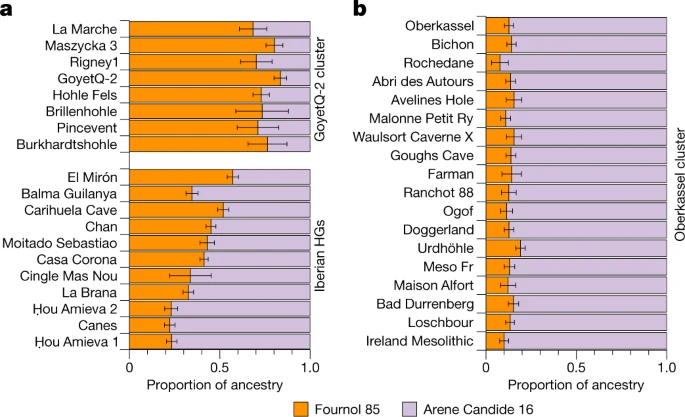 a,b, The ancestries of individuals in the GoyetQ-2 cluster and Iberian hunter-gatherers (HGs) (a) and individuals in the Oberkassel cluster (b) were modelled using qpAdm, with Fournol 85 and Arene Candide 16 representing the Fournol and Villabruna ancestries, respectively. The length of the colour bar shows the proportion of each ancestry. The error bar shows the s.e.m. of estimates from 5-cM-block jackknife analysis. Details of the modelling are provided in Supplementary Data 3.C. Our analyses demonstrate that the Fournol cluster is a better source for Magdalenian-associated genomes than Goyet Q116-1. Therefore, most of the ancestry found in these post-LGM individuals probably traced back to Gravettian-associated groups from western and southwestern Europe. The genetic affinity to the Villabruna ancestry is present in El Mirón and in Magdalenian-associated individuals from western and central Europe. This suggests that genetic links between southern and southwestern European hunter-gatherers around the time of the LGM extended north of the Pyrenees. The resulting GoyetQ2 cluster includes individuals spanning from western France to Poland in the period between 18 and 15 ka. Therefore, contrary to previous suggestions34, this demonstrates that the post-LGM diffusion of the Magdalenian was indeed associated with northward and northeastward population expansions from western Europe35. |
|
|
|
Post by Admin on Mar 13, 2023 19:47:44 GMT
Post-14 ka to Neolithic Previous studies have shown that two main hunter-gatherer ancestries were predominant across most parts of Europe after around 14 ka—that is, the western hunter-gatherer (WHG) ancestry, related to the Villabruna cluster, and the eastern hunter-gatherer (EHG) ancestry, showing affinity to both the Villabruna and the ancient north Eurasian (ANE) ancestry found in Upper Palaeolithic Siberian individuals4,36. Hunter-gatherers carrying an admixed WHG/EHG genetic profile have been sequenced from various regions of northern and eastern Europe, raising the question of how these two types of ancestries formed and interacted with each other through time and space37,38,39,40. In the MDS plot (Fig. 1c) and a west Eurasian principal component analysis (PCA) (Extended Data Fig. 6 and Supplementary Fig. 14), most post-14 ka individuals from western and central Europe fall close to the WHG cluster and those from eastern Europe close to the EHG cluster, whereas the Tutkaul 1 individual from central Asia falls close to the ANE-related group. The two 14 ka Oberkassel individuals mark the earliest presence of WHG ancestry north of the Alps, which we therefore rename the Oberkassel cluster (hereafter, Oberkassel cluster or ancestry), using the name of the oldest reported individual to date carrying such ancestry with more than one-fold coverage, for consistency4. On the basis of f4-statistics, we find that individuals assigned to the Oberkassel cluster are closer to the Arene Candide 16 genome than any other Epigravettian-associated group from Italy (Supplementary Data 2.F). Moreover, the Oberkassel cluster carries both Villabruna ancestry and a contribution from GoyetQ2 ancestry (Supplementary Data 2.J). This was confirmed with qpAdm, in which we could model all individuals from the Oberkassel cluster as a broadly constant mixture of approximately 75% Arene Candide 16 and 25% Goyet Q-2 (or 90% Arene Candide 16 and 10% Fournol 85) (Fig. 4b and Supplementary Data 3.C). The observation that post-14 ka individuals from western and central Europe and also from Britain41 carry a homogeneous genetic makeup instead of displaying repeated local admixtures with GoyetQ2 ancestry implies that the Oberkassel-ancestry profile was already largely formed before its dispersal. This is in sharp contrast to the genetic history of Iberian hunter-gatherers, where the spread of the Villabruna/Oberkassel ancestry involved multiple local admixture events with groups carrying high proportions of GoyetQ2 ancestry14 (Fig. 4 and Supplementary Data 3.C). The long-lasting genetic continuity in Iberia is also reflected in the preservation until the Mesolithic of Y-chromosome haplogroup C, which was predominant in pre-LGM groups but rarely found after the LGM in other parts of Europe (Extended Data Figs. 1 and 2). Using f4-statistics and qpAdm, we confirm that EHG populations in eastern Europe are a mixture of Villabruna/Oberkassel and ANE ancestries (Supplementary Information, section 11 and Supplementary Data 2.K). F4-statistics also show that the approximately 8.2 ka Yuzhniy Oleniy Ostrov group from Karelia in western Russia formed by 19 genomes has comparable or lower affinity to Villabruna ancestry than all the other EHG groups (Supplementary Data 2.K). The oldest individual revealing an indistinguishable genetic profile from the Yuzhniy Oleniy Ostrov group is the 11 ka Sidelkino individual from Samara in western Russia42. For consistency with the previously discussed nomenclature, we rename the EHG ancestry as the Sidelkino cluster (hereafter, Sidelkino cluster or ancestry). The genetic distinction between the Oberkassel and Sidelkino clusters is also clearly noticeable in the diversity of uniparentally inherited markers, as the Oberkassel cluster is dominated by mtDNA haplogroup U5 and Y-chromosome haplogroup I, whereas individuals from the Sidelkino cluster show a higher frequency of mtDNA haplogroups U2, U4 and R1b, and carry uniquely Y-chromosome haplogroups Q, R and J (Extended Data Figs. 1 and 2). We then attempted to model 250 published and newly reported hunter-gatherers dated to 14–5 ka using qpAdm as a mixture of Oberkassel, Sidelkino, GoyetQ2 ancestries, and an ancestry maximized in Anatolian Neolithic farmers (ANF), as a considerable portion of the sequenced hunter-gatherer genomes date after around 8 ka, when ANF ancestry started spreading across Europe. Our results show that the contact zone and the admixture patterns between the Oberkassel and Sidelkino ancestries changed over time (Fig. 5). Between 14 and 8 ka, all hunter-gatherers in western and central Europe carried only Oberkassel ancestry, with no detectable contribution from the Sidelkino cluster. Further north and east, individuals from the Baltics (Baltic HG), Scandinavia (SHG), the Balkans (Iron Gates HG) and Ukraine (Ukraine HG) already carried an Oberkassel/Sidelkino admixed ancestry38,40 before 8 ka. In addition, those groups also carry affinity to ANF suggesting more complex genetic processes behind their demographic history16. Moreover, two of the oldest published groups from western Russia belonging to the Sidelkino cluster—Peschanitsa (13 ka)43 and the newly reported Minino individuals (11 ka)—showed extra affinity to the Oberkassel cluster, possibly owing to variability in this ancestry proportion during the initial formation phase of the Sidelkino-ancestry profile. Using DATES software, we estimated the admixture between Villabruna/Oberkassel and ANE ancestries in these old Sidelkino-cluster-related individuals to around 15–13 ka (Extended Data Fig. 7 and Supplementary Table 3), which coincides roughly with the first appearance of the Oberkassel ancestry in central Europe. This raises the possibility that the replacement by the Oberkassel cluster and the formation of the Sidelkino cluster might have been the result of population expansions influenced by the abrupt warming during the Bølling–Allerød interstadial4,24. Fig. 5: Ancestry modelling of hunter-gatherers from 14–5.2 ka and their allele frequencies on phenotypic SNPs. 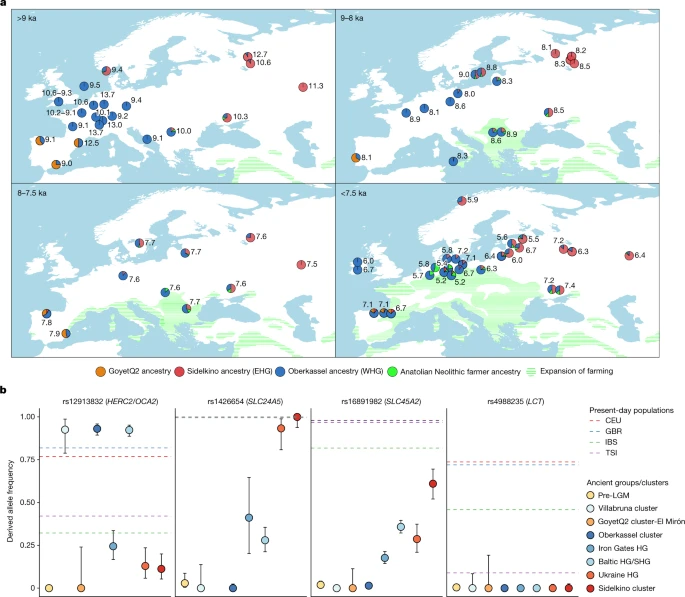 a, The genetic ancestry of hunter-gatherers dated between 14 ka and 5.2 ka modelled using qpAdm, with Oberkassel, Yuzhniy Oleniy Ostrov, Goyet Q-2 and Neolithic farmers from present-day Turkey (Barcın, Menteşe and Boncuklu sites) representing Oberkassel (WHG) (blue), Sidelkino (EHG) (red), GoyetQ2 (orange) and Anatolian Neolithic farmer (green) ancestries, respectively. The average calibrated date is shown, with pie charts indicating the estimated proportion of ancestry for each group or individual. Details of the modelling are provided in Supplementary Data 3.E,F. The expansion of farming by 9, 8, 7.5 and 7 ka is shown as green shades. Adapted from doi.org/10.5281/zenodo.5903165 (CC BY 4.0). b, Allele frequencies of different hunter-gatherer groups (coloured dots) on four SNPs related to skin colour (SLC24A5 and SLC45A2), eye colour (HERC2/OCA2) and lactase persistence (LCT). Dots are maximum likelihood estimates and error bars show 95% confidence intervals of the derived allele frequencies (n, the number of individuals in each group, is provided in Supplementary Data 3.G). Dashed lines show the frequencies estimated for the indicated present-day 1000 Genomes Project populations (CEU, Utah residents of northern and western European ancestry; GBR, British; IBS, Spanish; TSI, Tuscan )37. Details on the allele frequency estimates are provided in Supplementary Information, section 12, Supplementary Figs. 27 and 28 and Supplementary Data 3.G. From around 8 ka, we begin to observe admixture events with Sidelkino ancestry in central Europe. This is first detected in an individual from Gross Fredenwalde in northeastern Germany and reaches around 10% in most European hunter-gatherer individuals thereafter (Extended Data Fig. 8). Soon after 8 ka, Sidelkino ancestry was absent in eastern Spain but it had already reached northern Iberia alongside an increase in Oberkassel ancestry (Fig. 5). Conversely, additional Oberkassel ancestry is identified in eastern Europe by at least 7.5 ka in newly generated genomes from Minino I and Yazykovo from the upper Volga region, whereas a 1,000-years-older individual from Minino I did not have this genetic component. Considering a freshwater reservoir signal in the upper Volga region making radiocarbon dates on human remains appear up to about 500 years older than their true age44, there could be an interval of more than 1,000 years between the first evidence of admixture in central European hunter-gatherers with Sidelkino ancestry and eastern European hunter-gatherers with Oberkassel ancestry. However, additional genomes intermediate in time and space are needed to assess whether those two admixture events were independent or part of a common demographic process. After 7.5 ka, as ANF ancestry had reached regions north of the Alps, individuals carrying a hunter-gatherer genetic profile were primarily restricted to the northern fringes of Europe (Fig. 5). In this period, the Oberkassel-ancestry admixture spread further east, reaching Samara by around 6.5 ka, and an increase in Sidelkino ancestry was detected in hunter-gatherers from the Baltic region, which was previously associated with the transition from the Narva culture to the Comb Ceramic culture38,39 (Extended Data Fig. 8). In central Europe, admixture with ANF ancestry became highly common but not ubiquitous, indicating the co-existence of hunter-gatherer and farmer societies without admixing for several hundred years. The youngest individual carrying large portions of hunter-gatherer ancestry in the analysed dataset is from Ostorf in northern Germany, dated to around 5.2 ka (>90% Oberkassel cluster plus Sidelkino-cluster components) (Supplementary Data 3.F). Individuals at this site might mark one of the last occurrences of such high levels of hunter-gatherer-related ancestries, just centuries before the emerging European Bronze Age. On the basis of PCA and outgroup f3-statistics, the Neolithic Tutkaul 1 individual from Tajikistan is closely related to Upper Palaeolithic individuals from south-central Siberia (Afontova Gora 3 (AG3) and Mal’ta 1), and roughly contemporaneous West Siberian hunter-gatherers (Tyumen and Sosnoviy), both carrying high proportions of ANE ancestry45 (Fig. 1c and Extended Data Fig. 6). We tested the affinity of Tutkaul 1 to worldwide ancient and modern populations relative to AG3. Contrary to West Siberian hunter-gatherers, Tutkaul 1 does not carry an extra eastern Eurasian ancestry, but shows affinity to Iranian Neolithic farmers and some younger populations from Iran and the Turan region (Supplementary Data 2.L). Conversely, individuals in the Sidelkino cluster are genetically closer to AG3 than Tutkaul 1. This suggests that the newly reported Neolithic individual from central Asia carries an ancestry that might be a good proxy for the ANE-related contribution to Iran and the Turan region45 from around 5.5 ka but not to roughly contemporaneous hunter-gatherers from eastern Europe. In sum, we describe the formation and interaction between the Oberkassel and Sidelkino clusters, the two main hunter-gatherer ancestries present in Europe from 14 ka onwards. The genomic similarity of the Oberkassel cluster to Arene Candide 16 in northwestern Italy might imply that Epigravettian-associated ancestry spread from the south to central Europe passing through the western side of the Alpine region. The Sidelkino ancestry also emerged around 14 ka with its first direct evidence in eastern Europe43 dated to 13 ka. The increasing level of admixture between distinct hunter-gatherer populations from around 8 ka onwards indicates an intensified mobility of those forager groups. This might have been in part triggered by the concomitant expansion of Neolithic farmers across Europe and/or by environmental factors, such as the climatic event around 8.2 ka, the largest abrupt cooling in the northern hemisphere during the Holocene epoch46,47. |
|
|
|
Post by Admin on Mar 15, 2023 18:30:36 GMT
Discussion and conclusions The data generated in this study enabled us to investigate genomic transformations of and interactions between Eurasian hunter-gatherers at high resolution (Extended Data Fig. 9). We provide five novel insights into the genomic history of hunter-gatherer populations over a time span of 30,000 years from the Upper Palaeolithic to the Neolithic. 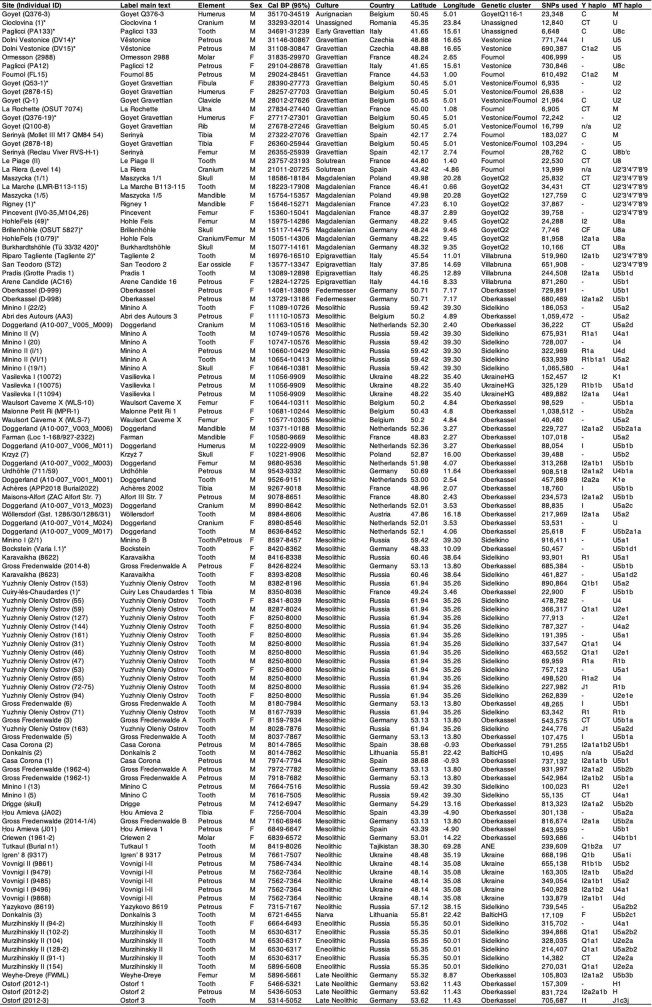 Extended Data Table 1 Summary statistics for individuals with newly reported genomic data First, we show that individuals associated with the Gravettian culture across Europe were not a biologically homogeneous population. Culturally, however, we see both widespread general tendencies, such as weaponry and some portable art48, and other aspects that have a more regional character, such as mortuary practices (Supplementary Information, section 13), various originalities in lithic and hard organic materials tool kits and adornments20,21. The ancestry found in individuals associated with the preceding Aurignacian culture from central Europe (GoyetQ116-1 ancestry) gave rise to Gravettian-associated individuals from western and southwestern Europe. This derived ancestry—the Fournol cluster—survived during the LGM in Solutrean-associated individuals, possibly within the Franco-Cantabrian climatic refugium25, leading to later populations associated with the Magdalenian culture (GoyetQ2 cluster and El Mirón). Conversely, the ancestry found in pre-30 ka eastern European individuals (Kostenki cluster and Sunghir group) contributed to Gravettian-associated individuals from central and southern Europe (Věstonice cluster), the latter without descendants retrieved in post-LGM populations from those regions. Second, the ancestry of individuals associated with the Epigravettian culture (Villabruna cluster), which was found to genetically connect European and Near Eastern hunter-gatherers, reached southern Europe well before the transition between the Early and Late Epigravettian4,15 and possibly as early as the Gravettian–Epigravettian transition. A phylogeographic reconstruction of different lineages carrying this ancestry further suggests its entry point into northeastern Italy from the Balkans followed by a north-to-south expansion into the Italian peninsula alongside a population decline through sequential bottlenecks. Third, Magdalenian-associated individuals not only from Iberia but also from the rest of Europe carry Epigravettian-associated ancestry (Villabruna cluster). Genetic analyses of western European individuals associated with the preceding Badegoulian culture might provide clues on the processes that led to the formation of the GoyetQ2 cluster. As inferred from the archaeological record35, the spread of the Magdalenian across Europe is linked to southwestern to northern and northeastern post-LGM population expansions and not to movements from southeastern refugia34. Fourth, we extend the finding of a large-scale genetic turnover as early as 14 ka in central and western European hunter-gatherers associated with multiple techno-complexes—Federmesser, Azilian and other Final Palaeolithic groups4—despite considerable technological continuity with the preceding late Magdalenian. This broadly distributed ancestry (the Oberkassel cluster (also known as WHG)) is most closely related to an Epigravettian-associated individual from northwestern Italy, suggesting that its expansion into continental Europe might have started from the west—and not the east—side of the Alps. Moreover, the almost complete genetic replacement of the Magdalenian-associated gene pool raises the hypothesis that parts of Europe were differentially populated during the abrupt climatic variation starting around 14.7 ka with the Bølling–Allerød warming period, creating areas where southern European populations could expand. This might also explain the genetic uniformity of the Oberkassel cluster across large parts of western Eurasia but genomic data from between 15 and 14 ka is needed to understand the exact dynamics of this turnover. Fifth, the Oberkassel ancestry in western and central Europe and the Sidelkino ancestry in eastern Europe remained largely isolated for almost 6,000 years until genetic interactions were first observed—around 8 ka in northeastern Germany, possibly associated with cultural exchanges along the Baltics49 and around 7.5 ka in the upper Volga region, possibly linked to the spread of pottery in the region50. In conclusion, our study reveals that western and southwestern Europe served as climatic refugia for the persistence of human groups during the coldest phase of the last Ice Age whereas populations in the Italian peninsula and the eastern European plain were genetically overturned, challenging the role of these regions as glacial refugia for humans. The incoming Villabruna ancestry later became the most widespread hunter-gatherer ancestry across Europe. Further palaeogenomic studies on Upper Palaeolithic individuals from the Balkans will be essential for understanding whether southeastern Europe represents the source of the Villabruna ancestry and a climatic refugium for human populations during the LGM. Note added in proof: A companion paper51 describes genome-wide data of a 23,000-year-old Solutrean-associated individual from southern Iberia that extend the evidence of genetic continuity across the LGM in southwestern Europe. www.nature.com/articles/s41586-023-05726-0#Fig6 |
|

