

Post by Admin on May 5, 2014 2:12:03 GMT
Villa and Roebroeks say that the past misrepresentation of Neanderthals’ cognitive ability may be linked to the tendency of researchers to compare Neanderthals, who lived in the Middle Paleolithic, to modern humans living during the more recent Upper Paleolithic period, when leaps in technology were being made. Although many still search for a simple explanation and like to attribute the Neanderthal demise to a single factor, such as cognitive or technological inferiority, archaeology shows that there is no support for such interpretations, the authors said.
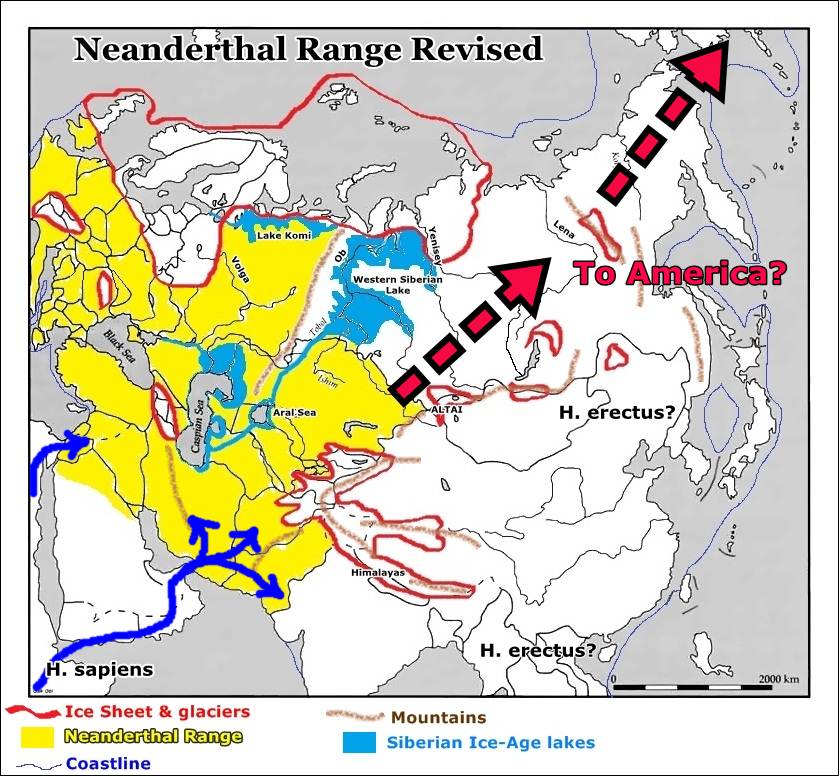
We conclude that all the “archaeology-based” explanations for the demise of the Neandertals reviewed here (Table 1, Text S1, Hypotheses 1–11) are flawed. They were based on much less data than we have available today and were at least in part the result of a long tradition of thinking in terms of Neandertals-AMH dichotomies, steered by overstressing developments within the Upper Paleolithic of Europe, the record of which has become almost like a yardstick for modern human behavior (Text S2).
While the debate about AMH dispersal times and routes out of Africa is intense, based on a range of archaeological as well as genetic data, the archaeological record from the various continents does not provide strong support for any of the suggested routes nor any of the suggested factors in the demise of the Neandertals. The very fact that the migration time estimates vary so widely suggests that we simply have no solid data; perhaps there was more than one migration event (in addition to the Last Interglacial limited expansion in the Levant), and in all probability the migrating groups did not have a strong cultural homogeneity. This may explain why we do not see clear archaeological signatures for AMH on the move.
Interestingly, the widely accepted date of 60 or 50 ka for the modern human expansion into Eurasia (following the earlier short-lived exodus in the Levant documented at Skuhl and Qafzeh) would rule out South Africa as the location for source populations for two reasons: (i) by 60 ka the HP tradition of backed tools made on blades and bladelets produced by soft stone hammer (supposedly associated with the AMH expansion) had given way to the post-HP assemblages characterized by a variety of flake tools and blades produced by hard hammer percussion but without backed blades [94], [103], [105], [116]; (ii) the Still Bay and HP populations were not larger than other MSA populations and might even have been smaller, thus excluding population pressure as the prime mover of the migration [117]. According to Klein [11] the Out of Africa expansion was underlain by a neural mutation that promoted the final development of the modern human brain. Direct evidence for this hypothesis may come from comparisons of Neandertal and modern human genomes.

In the recent past, much debate has been generated from the observation that Neandertals began to produce a richer archaeological record, including bone tools, personal ornaments and use of manganese and ochre, at the time when AMH started colonizing Europe. Some interpreted this change in the record as the result of Neandertal absorption of ideas and techniques from the incoming AMH. After having produced a rather monotonous record for almost 300,000 years, an independent invention of these new items just at the time of the arrival of AMH would have to be seen as an “impossible coincidence” [28]. However, as reviewed here, use of ochre, of personal ornaments, production of specialized bone tools and complex hafting techniques were part of the Neandertal repertoire already before the arrival of AMH in western Eurasia.
The present review also suggests that some of the innovative technologies of the Protoaurignacian and of the Aurignacian may have developed out of a Middle Paleolithic base (for a similar viewpoint, see [118]). Some components that occur sporadically or episodically in Neandertal and late MSA assemblages become much more common later, like pigment use, symbolic objects, extensive transport of raw materials and even specialized bone tools [79]. The same goes for another element, the intentional production of bladelets (<4 cm in length) from bladelet cores. Bladelets have been considered a discriminant factor between the Upper and Middle Paleolithic and therefore between AMH and Neandertals [119]. Production of bladelets has been securely identified in French Mousterian assemblages, e.g. at Combe Grenal (layers 30–29 and layers 16 and 14), Champ Grand and Grotte Mandrin, and in Spain at sites such as El Castillo and Cueva Morin [120], [121]. All these assemblages belong to the final Mousterian, with the exception of Combe Grenal and Grotte Mandrin; at the latter site, a layer with blades, bladelets and microlithic points is overlain by five layers with flake-based Mousterian assemblages [121]. At Combe Grenal layers 29–30 have an estimated age of late MIS 4, i.e. around 60 ka. Bladelets and bladelet cores are not abundant (5% of the assemblage at Combe Grenal layers 29–30), yet they show that Neandertals, like late MSA humans and the makers of the Protoaurignacian, mastered the technology of bladelet production, albeit using methods different from the HP small blade technology. It is their frequency, not cognition or technical competence, that distinguishes AMH bladelet production from that of Neandertals [120]. The techniques and methods of bladelet making in the Mousterian are different from those of the Protoaurignacian, just as the kind of possible symbolic objects are also different (use of raptor claws; on perforated or grooved animal teeth (see Text SI, Hypothesis 1). Perhaps the nature of the contacts should be seen in terms of diffusion of ideas rather than as face to face interaction and the copying of specific objects [122]. The occurrence of Dufour bladelets (often used as projectile elements in the Aurignacian and the Protoaurignacian) with very specific techniques of manufacture in the Châtelperronian of Quincay is interpreted in a similar way, as a form of low-degree social interaction between Neandertals and modern humans [123].
The Date of the Demise
Various new dates support the idea of some chronological overlap between AMH and Neandertals, which may have enabled interbreeding and cultural interaction in western Europe: AMS dates on ultrafiltered bone collagen from the Châtelperronian layers X and IX of Grotte du Renne at Arcy, c. 44 to 41 kyr cal BP; the date of the Saint-Césaire Neandertal at 41.9−40.6 kyr cal BP [63]; the fact that the Protoaurignacian at the Italian sites of Castelcivita and Serino is overlain by the Campanian Ignimbrite tephra, dated to 39.28±0.11 ka by 40Ar/39Ar [124]; the modeled age ranges of c. 41.5−39.9 kyr cal BP of several radiocarbon-dated Proto-Aurignacian sites [95]; the date of the Oase 2 early modern cranium at c. 40 ka [125]; the AMS dates for the Neandertal child from Spy cave (Belgium), 36,870 to 38,494 and 37,297 to 40,490 cal BP [126]; the AMS dates for the Vindija (Croatia) Neandertal remains at c. 38 kyr cal BP [127], [128]; the 37.4 ka cal BP date for the final Mousterian level of Cueva Antón in southeastern Spain [75]. Even if we do not consider dates judged by some as controversial such as (i) the AMS dates on shell beads for the layer containing the modern human teeth at Grotta del Cavallo at 45.010−43,380 cal BP [5, contra 2]; (ii) the dates for the Kent’s Cavern modern human maxilla [3, contra 2, 129]; and (iii) the dates for the Aurignacian at Geissenklösterle at c. 42 kyr cal BP [4, contra 2], some millennia of overlap are indicated The latest Neandertal currently known from the Levant is the adult male skeleton from Amud Cave (Israel) with an ESR date of 53±8 ka on tooth enamel [130].
Interbreeding and Assimilation
For some authors replacement and supposedly rapid extinction of Neandertals can be explained only in terms of substantial cognitive, technological and demographic differences between the Neandertals and AMH [42], [131]. But, as we tried to show here, the Neandertal archaeological record was not different enough to explain their demise in terms of inferiority in archaeologically visible domains. Thus, if Neandertals were not technologically and cognitively “disadvantaged”, how can we explain that they did not survive?

Some modern human-like anatomical characteristics are said to occur in late Neandertal fossils (as in the Vindja, St. Césaire and Riparo Mezzena late Neandertals [132], [133] and refs therein) and vice versa some Neandertal features are present in early specimens of modern humans in Europe [134], [135] supporting a hypothesis of some degree of admixture between the two groups. However, until recently the morphological evidence of admixture was often dismissed. In 2010 a draft sequence of the Neandertal nuclear DNA provided clear evidence of interbreeding between Neandertals and modern humans [48], estimating that Neandertal inheritance makes up 1–4% of the genomes of people outside of Africa. A revised estimate based on a high-coverage sequence of a Neandertal from the Altai Mountains now suggests 1.5–2.1% [49]. Genes of Neandertals may have been favored through natural selection, and possibly played a role in the development of the immune system of modern humans [136] or in UV-light adaptations [137]. According to [138] gene flow from Neandertals to modern humans occurred between 47,000 and 65,000 years ago, and most likely happened at the time when Neandertals and modern humans encountered each other in Europe and the Middle East around 50,000 years ago.
In sum, interbreeding and assimilation, the tenants of a model first proposed by Fred Smith [139] are now supported by genetic data [134], [140]. It can be argued that the level of interbreeding may have been too limited to support an assimilation scenario. An interestingly parallel to this complex situation can be found in another “revolution”, the so-called Neolithic Revolution [43], [141], which does not feature explanations in terms of “cognitive” differences. The first farmers swept into Europe from the Near East at about 7500y BP displacing the local Late Mesolithic hunter-gatherers. But the Mesolithic hunter-gatherers, who cannot be described as cognitively inferior, were not submerged by hordes of farmers. Farmers and foragers coexisted for thousands of years in NW Europe; in Central Europe local hunter-gatherers adopted farming but in southern Scandinavia local foragers retained the Mesolithic lifestyle for c. 1500 years after farming arrived in Central Europe [142], [143]. Cultural contact is suggested by clear continuities in flint technology between the Mesolithic and early Neolithic in the region. After a very complex pattern of expansions and genetic shifts of the last 8,000 years the hunter-gatherer mitochondrial DNA haplogroups form 16% of the present-day Central European genetic composition [143]. It would take at least one millennium between the first arrival of immigrants and a notable increase in their population size.
The original Neandertal contribution to modern human biology may have been larger 40,000 years ago - equivalent to 2000 generations (with generation time at 20 years) – than estimates based on genomic regions of present-day humans suggest [144], [145]. Interbreeding of Neandertals and modern humans may have helped modern humans to adapt to non-African environments but also introduced alleles that were not tolerated and contributed to male hybrid sterility thus reducing the proportion of Neandertal ancestry of the period of contact to that seen today [144].
Mitochondrial genetic diversity of eight early modern European humans dated to ca 38,000 to 4,500 (14C cal BP, from Kostienki 14 to Őtzi The Iceman) is 1.5 times higher than that of five European Neandertals spanning the time to 38 to 70 ka [23], [146]. The high coverage genome of the Altai Neandertal [49] also suggests low genetic diversity which could indicate small population sizes (see Text S1 Hypothesis 8 for archaeological data). These genetic data suggest that differences in population sizes between the “resident” Neandertals and incoming AMH populations may have been a contributing factor in the absorption of Neandertal populations [23]. The momentous cultural changes that followed the arrival of AMH in Western Eurasia were not uniquely due to the residents’ cognitive or technological inferiority causing rapid and total replacement. The Neandertal demise appears to have resulted from a complex and protracted process [147] including multiple dynamic factors such as low population density, interbreeding with some cultural contact, possible male hybrid sterility and contraction in geographic distribution [148] followed by genetic swamping and assimilation by the increasing numbers of modern immigrants.
Conclusion
In a review of the MSA and Middle Paleolithic archaeological record we have shown that inferred markers of modern human cognitive and behavioral capacities have a greater time depth in the Middle Palaeolithic record than commonly acknowledged. We have found no data in support of the supposed technological, social and cognitive inferiority of Neandertals compared to their AMH contemporaries. The results of our study imply that single-factor explanations for the disappearance of the Neandertals are not warranted any more, and that their demise was clearly more complex than many archaeology-based scenarios of “cognitive inferiority” reviewed here seem to suggest. This has implications beyond the field of archaeology per se: archaeologists’ characterizations of Neandertals as cognitively inferior to modern humans [149] have created an interpretive framework within which subtle biological differences between Neandertals and modern humans tend to be overinterpreted (see for instance [150].
After 40,000 years and 2000 generations the Neandertal fraction in non-African modern human genomes still constitutes a substantial legacy from these ancient hominins who differed from contemporary AMHs in both geno- and phenotypes [151] but whose archeological record was not different enough to support the purported cognitive “gap” between them and their contemporary modern humans.
www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0096424
We conclude that all the “archaeology-based” explanations for the demise of the Neandertals reviewed here (Table 1, Text S1, Hypotheses 1–11) are flawed. They were based on much less data than we have available today and were at least in part the result of a long tradition of thinking in terms of Neandertals-AMH dichotomies, steered by overstressing developments within the Upper Paleolithic of Europe, the record of which has become almost like a yardstick for modern human behavior (Text S2).
While the debate about AMH dispersal times and routes out of Africa is intense, based on a range of archaeological as well as genetic data, the archaeological record from the various continents does not provide strong support for any of the suggested routes nor any of the suggested factors in the demise of the Neandertals. The very fact that the migration time estimates vary so widely suggests that we simply have no solid data; perhaps there was more than one migration event (in addition to the Last Interglacial limited expansion in the Levant), and in all probability the migrating groups did not have a strong cultural homogeneity. This may explain why we do not see clear archaeological signatures for AMH on the move.
Interestingly, the widely accepted date of 60 or 50 ka for the modern human expansion into Eurasia (following the earlier short-lived exodus in the Levant documented at Skuhl and Qafzeh) would rule out South Africa as the location for source populations for two reasons: (i) by 60 ka the HP tradition of backed tools made on blades and bladelets produced by soft stone hammer (supposedly associated with the AMH expansion) had given way to the post-HP assemblages characterized by a variety of flake tools and blades produced by hard hammer percussion but without backed blades [94], [103], [105], [116]; (ii) the Still Bay and HP populations were not larger than other MSA populations and might even have been smaller, thus excluding population pressure as the prime mover of the migration [117]. According to Klein [11] the Out of Africa expansion was underlain by a neural mutation that promoted the final development of the modern human brain. Direct evidence for this hypothesis may come from comparisons of Neandertal and modern human genomes.
In the recent past, much debate has been generated from the observation that Neandertals began to produce a richer archaeological record, including bone tools, personal ornaments and use of manganese and ochre, at the time when AMH started colonizing Europe. Some interpreted this change in the record as the result of Neandertal absorption of ideas and techniques from the incoming AMH. After having produced a rather monotonous record for almost 300,000 years, an independent invention of these new items just at the time of the arrival of AMH would have to be seen as an “impossible coincidence” [28]. However, as reviewed here, use of ochre, of personal ornaments, production of specialized bone tools and complex hafting techniques were part of the Neandertal repertoire already before the arrival of AMH in western Eurasia.
The present review also suggests that some of the innovative technologies of the Protoaurignacian and of the Aurignacian may have developed out of a Middle Paleolithic base (for a similar viewpoint, see [118]). Some components that occur sporadically or episodically in Neandertal and late MSA assemblages become much more common later, like pigment use, symbolic objects, extensive transport of raw materials and even specialized bone tools [79]. The same goes for another element, the intentional production of bladelets (<4 cm in length) from bladelet cores. Bladelets have been considered a discriminant factor between the Upper and Middle Paleolithic and therefore between AMH and Neandertals [119]. Production of bladelets has been securely identified in French Mousterian assemblages, e.g. at Combe Grenal (layers 30–29 and layers 16 and 14), Champ Grand and Grotte Mandrin, and in Spain at sites such as El Castillo and Cueva Morin [120], [121]. All these assemblages belong to the final Mousterian, with the exception of Combe Grenal and Grotte Mandrin; at the latter site, a layer with blades, bladelets and microlithic points is overlain by five layers with flake-based Mousterian assemblages [121]. At Combe Grenal layers 29–30 have an estimated age of late MIS 4, i.e. around 60 ka. Bladelets and bladelet cores are not abundant (5% of the assemblage at Combe Grenal layers 29–30), yet they show that Neandertals, like late MSA humans and the makers of the Protoaurignacian, mastered the technology of bladelet production, albeit using methods different from the HP small blade technology. It is their frequency, not cognition or technical competence, that distinguishes AMH bladelet production from that of Neandertals [120]. The techniques and methods of bladelet making in the Mousterian are different from those of the Protoaurignacian, just as the kind of possible symbolic objects are also different (use of raptor claws; on perforated or grooved animal teeth (see Text SI, Hypothesis 1). Perhaps the nature of the contacts should be seen in terms of diffusion of ideas rather than as face to face interaction and the copying of specific objects [122]. The occurrence of Dufour bladelets (often used as projectile elements in the Aurignacian and the Protoaurignacian) with very specific techniques of manufacture in the Châtelperronian of Quincay is interpreted in a similar way, as a form of low-degree social interaction between Neandertals and modern humans [123].
The Date of the Demise
Various new dates support the idea of some chronological overlap between AMH and Neandertals, which may have enabled interbreeding and cultural interaction in western Europe: AMS dates on ultrafiltered bone collagen from the Châtelperronian layers X and IX of Grotte du Renne at Arcy, c. 44 to 41 kyr cal BP; the date of the Saint-Césaire Neandertal at 41.9−40.6 kyr cal BP [63]; the fact that the Protoaurignacian at the Italian sites of Castelcivita and Serino is overlain by the Campanian Ignimbrite tephra, dated to 39.28±0.11 ka by 40Ar/39Ar [124]; the modeled age ranges of c. 41.5−39.9 kyr cal BP of several radiocarbon-dated Proto-Aurignacian sites [95]; the date of the Oase 2 early modern cranium at c. 40 ka [125]; the AMS dates for the Neandertal child from Spy cave (Belgium), 36,870 to 38,494 and 37,297 to 40,490 cal BP [126]; the AMS dates for the Vindija (Croatia) Neandertal remains at c. 38 kyr cal BP [127], [128]; the 37.4 ka cal BP date for the final Mousterian level of Cueva Antón in southeastern Spain [75]. Even if we do not consider dates judged by some as controversial such as (i) the AMS dates on shell beads for the layer containing the modern human teeth at Grotta del Cavallo at 45.010−43,380 cal BP [5, contra 2]; (ii) the dates for the Kent’s Cavern modern human maxilla [3, contra 2, 129]; and (iii) the dates for the Aurignacian at Geissenklösterle at c. 42 kyr cal BP [4, contra 2], some millennia of overlap are indicated The latest Neandertal currently known from the Levant is the adult male skeleton from Amud Cave (Israel) with an ESR date of 53±8 ka on tooth enamel [130].
Interbreeding and Assimilation
For some authors replacement and supposedly rapid extinction of Neandertals can be explained only in terms of substantial cognitive, technological and demographic differences between the Neandertals and AMH [42], [131]. But, as we tried to show here, the Neandertal archaeological record was not different enough to explain their demise in terms of inferiority in archaeologically visible domains. Thus, if Neandertals were not technologically and cognitively “disadvantaged”, how can we explain that they did not survive?
Some modern human-like anatomical characteristics are said to occur in late Neandertal fossils (as in the Vindja, St. Césaire and Riparo Mezzena late Neandertals [132], [133] and refs therein) and vice versa some Neandertal features are present in early specimens of modern humans in Europe [134], [135] supporting a hypothesis of some degree of admixture between the two groups. However, until recently the morphological evidence of admixture was often dismissed. In 2010 a draft sequence of the Neandertal nuclear DNA provided clear evidence of interbreeding between Neandertals and modern humans [48], estimating that Neandertal inheritance makes up 1–4% of the genomes of people outside of Africa. A revised estimate based on a high-coverage sequence of a Neandertal from the Altai Mountains now suggests 1.5–2.1% [49]. Genes of Neandertals may have been favored through natural selection, and possibly played a role in the development of the immune system of modern humans [136] or in UV-light adaptations [137]. According to [138] gene flow from Neandertals to modern humans occurred between 47,000 and 65,000 years ago, and most likely happened at the time when Neandertals and modern humans encountered each other in Europe and the Middle East around 50,000 years ago.
In sum, interbreeding and assimilation, the tenants of a model first proposed by Fred Smith [139] are now supported by genetic data [134], [140]. It can be argued that the level of interbreeding may have been too limited to support an assimilation scenario. An interestingly parallel to this complex situation can be found in another “revolution”, the so-called Neolithic Revolution [43], [141], which does not feature explanations in terms of “cognitive” differences. The first farmers swept into Europe from the Near East at about 7500y BP displacing the local Late Mesolithic hunter-gatherers. But the Mesolithic hunter-gatherers, who cannot be described as cognitively inferior, were not submerged by hordes of farmers. Farmers and foragers coexisted for thousands of years in NW Europe; in Central Europe local hunter-gatherers adopted farming but in southern Scandinavia local foragers retained the Mesolithic lifestyle for c. 1500 years after farming arrived in Central Europe [142], [143]. Cultural contact is suggested by clear continuities in flint technology between the Mesolithic and early Neolithic in the region. After a very complex pattern of expansions and genetic shifts of the last 8,000 years the hunter-gatherer mitochondrial DNA haplogroups form 16% of the present-day Central European genetic composition [143]. It would take at least one millennium between the first arrival of immigrants and a notable increase in their population size.
The original Neandertal contribution to modern human biology may have been larger 40,000 years ago - equivalent to 2000 generations (with generation time at 20 years) – than estimates based on genomic regions of present-day humans suggest [144], [145]. Interbreeding of Neandertals and modern humans may have helped modern humans to adapt to non-African environments but also introduced alleles that were not tolerated and contributed to male hybrid sterility thus reducing the proportion of Neandertal ancestry of the period of contact to that seen today [144].
Mitochondrial genetic diversity of eight early modern European humans dated to ca 38,000 to 4,500 (14C cal BP, from Kostienki 14 to Őtzi The Iceman) is 1.5 times higher than that of five European Neandertals spanning the time to 38 to 70 ka [23], [146]. The high coverage genome of the Altai Neandertal [49] also suggests low genetic diversity which could indicate small population sizes (see Text S1 Hypothesis 8 for archaeological data). These genetic data suggest that differences in population sizes between the “resident” Neandertals and incoming AMH populations may have been a contributing factor in the absorption of Neandertal populations [23]. The momentous cultural changes that followed the arrival of AMH in Western Eurasia were not uniquely due to the residents’ cognitive or technological inferiority causing rapid and total replacement. The Neandertal demise appears to have resulted from a complex and protracted process [147] including multiple dynamic factors such as low population density, interbreeding with some cultural contact, possible male hybrid sterility and contraction in geographic distribution [148] followed by genetic swamping and assimilation by the increasing numbers of modern immigrants.
Conclusion
In a review of the MSA and Middle Paleolithic archaeological record we have shown that inferred markers of modern human cognitive and behavioral capacities have a greater time depth in the Middle Palaeolithic record than commonly acknowledged. We have found no data in support of the supposed technological, social and cognitive inferiority of Neandertals compared to their AMH contemporaries. The results of our study imply that single-factor explanations for the disappearance of the Neandertals are not warranted any more, and that their demise was clearly more complex than many archaeology-based scenarios of “cognitive inferiority” reviewed here seem to suggest. This has implications beyond the field of archaeology per se: archaeologists’ characterizations of Neandertals as cognitively inferior to modern humans [149] have created an interpretive framework within which subtle biological differences between Neandertals and modern humans tend to be overinterpreted (see for instance [150].
After 40,000 years and 2000 generations the Neandertal fraction in non-African modern human genomes still constitutes a substantial legacy from these ancient hominins who differed from contemporary AMHs in both geno- and phenotypes [151] but whose archeological record was not different enough to support the purported cognitive “gap” between them and their contemporary modern humans.
www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0096424










