|
|
Post by Admin on Jul 11, 2022 23:55:44 GMT
To explore this further, we used f4-admixture statistics of the form f4(Chimp, Sicily LM HG; Anatolia EN Barcin, Sicily EN) to test whether the Sicily EN farmers retained an excess of shared alleles with the preceding Sicily LM HGs, when using Anatolia EN Barcin as a baseline for EEF ancestry. The statistic was significantly positive, suggesting excess local HG ancestry in Sicily EN farmers (z = 5.051, Data S3.2). Indeed, with qpAdm-based admixture models, we estimate that local Sicilian LM HGs contributed 7.5 ± 0.9% of the ancestry to the Sicilian EN farmer gene pool (Figure 3A, Data S3.3). Of note, the local HG admixture signal is significantly lower when compared to previously published Sicily MN farmers from Fossato di Stretto Partanna (Fernandes et al., 2020) (f4(Chimp, Sicily LM HG; Sicily EN, and Sicily MN): z = 5.381, Data S3.2), which carried an estimated 11.9 ± 0.9% local Sicily LM HG ancestry (Data S3.3). Overall, this suggests a subtle, yet detectable, contribution and potential resurgence of local HG ancestry in Sicilian farmers during the Neolithic. Importantly, the published Sicily MN farmers are contemporaneous with the youngest farmer individual UZZ61 (6,830–6,660 cal BP) in our Sicily EN group, which indicates that the resurgence may have taken place elsewhere in Sicily (e.g., at Fossato di Stretto Partanna) by the mid-seventh-millenium BP, but not yet in G. dell’Uzzo.
The Early Bronze Age individual UZZ57 shows a shift in PC space toward individuals that are associated with the Late Neolithic Bell Beaker phenomenon and EBA groups that carry steppe-related ancestry (Figure 1C), similar to what has been observed in other EBA individuals from Sicily (Fernandes et al., 2020). This is also confirmed by qpAdm, in which individual UZZ57 could be modeled as a two-way mixture between local Sicily EN ancestry and ‘steppe ancestry’ represented by Yamnaya Samara (Figure 3A). The steppe ancestry contribution in UZZ57 was estimated to 21.0 ± 3.5%, which falls within the range of other published Sicily EBA groups (Data S3.4).
Did Sicilian Late Mesolithic foragers adopt some aspects of early farming?
To shed further light on the processes underlying the Mesolithic-Neolithic transition in Sicily, we jointly analyzed the stable isotope data for dietary inference (δ13C/δ15N) and the ancestry profile for each individual in our time transect (Data S5.1, Figure S2, Table S3). We find that individuals from different time periods carrying different genetic ancestries also consumed isotopically different diets (Figure 3B). The isotopic data show that the Early Mesolithic HGs from G. dell’Uzzo relied mainly on hunting terrestrial game with substantial contributions of plant foods but limited consumption of marine resources (Mannino et al., 2015). In contrast, the diet of the Sicily LM HGs carrying EHG ancestry was characterized by a significantly higher intake of marine-based protein. The isotopic values for the Sicilian EN farmers are congruent with them having a terrestrial-based farming diet. Overall, the isotopic and ancestry profiles per individual show that diets correspond broadly with the assigned genomic cluster and attested subsistence per archaeological time period.
However, two individuals (UZZ71, UZZ88) chronologically overlap with the period when the Impressa Ware made its appearance at the site (Mannino et al., 2015). The two oldest individuals (UZZ71 and UZZ88), genetic females dated to ∼7,960–7,790 cal BP, fall fully within the genomic diversity of the Late Mesolithic HGs associated with the Castelnovian sensu lato, despite postdating them by ∼200 years (Figure 1C, Data S5.1). Both these individuals show isotope values that are different from the preceding Late Mesolithic HGs as well as from the later Sicily EN farmers associated with Stentinello/Kronio pottery (Figure 3B, Data S5.1). The dietary profile of individual UZZ71 (δ13C = −18.9‰, δ15N = 14.5‰) indicates an intake of freshwater protein, similarly to what has been reported for Mesolithic HG from the Iron Gates on the Balkan Peninsula (Bonsall et al., 2015; Borić and Price, 2013). On the other hand, individual UZZ88 shows an isotopic composition (δ13C = −19.2‰, δ15N = 7.1‰) that suggests a terrestrial-based farming diet with very low levels of animal protein consumption (EN farmers analyzed from G. dell’Uzzo): mean δ13C = −19.4 ± 0.5‰, mean δ15N = 9.1 ± 1.2‰). The observed combination of HG ancestry profile and terrestrial farming diet for UZZ88 can be explained in two ways: agro-pastoralism had strongly influenced local subsistence practices and/or some foragers had become part of the incoming farming groups. Conversely, UZZ77, an individual whose genetic makeup is typical of the incoming farmers, has a dietary profile (δ13C = −18.8‰, δ15N = 12.5‰) more similar to that of the Mesolithic terrestrially-based foragers. The combined genetic and isotopic data for UZZ88 and UZZ77 point to some degree of interaction between local hunter-gatherers and incoming farmers, which is also compatible with the small proportion of HG ancestry present in Sicily after the introduction of farming, as also attested at the MN site of Fossato di Stretto Partanna (Fernandes et al., 2020).
|
|
|
|
Post by Admin on Jul 12, 2022 15:44:30 GMT
Discussion The Apennine Peninsula (today’s Italy) has long been viewed as one of the refugia during the LGM (Feliner, 2011; Schmitt, 2007), ∼25,000–18,000 years ago, from where Europe was repopulated (Fu et al., 2016; Posth et al., 2016). The earliest evidence for the presence of Homo sapiens in Sicily dates to ∼19,000–18,000 cal BP, following the time when a land bridge connected the island to peninsular Italy (Antonioli et al., 2016; Di Maida et al., 2019). Although there are many sites in peninsular Italy and Sicily with evidence of Late Upper Paleolithic occupation, forager populations seemed to have decreased during the Mesolithic (Biagi, 2003), congruent with the observation that EM HGs from G. dell’Uzzo produced a lithic industry derived from the Epigravettian tradition (STAR Methods) (Lo Vetro and Martini, 2016). The profoundly reduced population genomic diversity (Figure S4), large quantity of short ROH tracts in the Sicily EM HGs (Figure S5), together with a high level of shared genetic drift with Mesolithic foragers from Continenza (Figure S3, Data S2.1), hint at a population bottleneck that affected foragers in Sicily and potentially also central Italy. Some Late Epigravettian sites in Sicily contain rock panels with engraved animal figures that are similar to those of the Franco-Cantabrian, including Magdalenian (Mussi, 2001). Here, we showed that compared to the ∼14,000 yBP Epigravettian-associated HG Villabruna in northern Italy, the Sicily EM HGs have a higher genetic affinity to Iberian HGs associated with the Magdalenian and Azilian, such as El Mirón and Balma Guilanyà (Data S2.2). Antonio et al. reported similar affinities for Mesolithic HGs from Continenza (Antonio et al., 2019). Compared to Villabruna, no extra Magdalenian-related ancestry has been found in Continenza or G. dell’Uzzo (Data S2.2) (Antonio et al., 2019), whereas Iberian HGs including El Mirón and Balma Guilanyà have been suggested to carry Epigravettian-related ancestry (Figure S6) (van de Loosdrecht, 2021; Villalba-Mouco et al., 2019). This suggests the Epigravettian-related ancestry in Iberian HGs was likely derived from southern/central Italy. Both archaeological and genomic results suggest a deep connection between the Epigravettian of southern Italy and the Magdalenian and Azilian in Iberia (van de Loosdrecht, 2021), which warrants further investigation. Compared to Sicily EM HGs, Sicily LM HGs derived ∼18–21% of their ancestry from an EHG-related source, providing evidence for shifting ancestry during the Mesolithic (Figure 3A). After the LGM, foragers associated with Epigravettian assemblages expanded in both Italy and southeastern Europe (Djindjian, 2016; Maier, 2015). Similar to southern Italy, the Balkans were also a glacial refugium. Currently, there are no genomes available for Epigravettian foragers from the Balkans. However, the Mesolithic HGs from this region are among the oldest HGs carrying EHG-related ancestry in southeastern Europe (Figure S6C) and suggest the Balkans as a candidate region for the excess EHG ancestry found in LM Sicily HGs. From the start of the Early Holocene, ∼11,700 yBP onward, an EHG/Anatolia HG (AHG) mixture can be found among HGs from Scandinavia, the Baltic, Ukraine, and the Balkans (Figure S6) (van de Loosdrecht, 2021). This underlines previous reports for a long-standing interaction sphere of HGs with EHG and AHG-related ancestry from northern and eastern Europe toward the Near East and the Caucasus (Feldman et al., 2019; Fu et al., 2016; Mathieson et al., 2018). This population may well have expanded into Sicily in the course of the westward shift of the blade-and-trapeze horizon into central and southern Europe, which seems to have begun after the climatic anomaly around 9,300 yBP (Gronenborn, 2017; van de Loosdrecht, 2021). In contrast, the ancient genome-wide data for the Sicily EN farmers point to a near-complete genetic turnover during the transition from foraging to farming. The preceding Late Mesolithic HGs have contributed at most ∼7% ancestry to the EN farmers from G. dell’Uzzo (Figure 3A). This indicates that the transition to agriculture involved population replacement of local foragers by early farmers to a large extent during the Early Neolithic, similar to previous results for early farmers from Ripabianca and Continenza in peninsular Italy and other regions in Europe (Antonio et al., 2019; Bramanti et al., 2009; Günther et al., 2015; Hofmanová et al., 2016; Kılınç et al., 2016; Lazaridis et al., 2014, Lazaridis et al., 2016, Lazaridis et al., 2017). However, the distinct diets and the intermediate 14C dates of a few individuals from the period around the beginning of the Neolithic provide tentative evidence that HGs and farmers initially exchanged their subsistence practices in Sicily, as was hypothesized by Tusa (Tusa, 1985, Tusa, 1996). We observed individuals with HG ancestry and a farming/fishing diet, while one individual with farmer ancestry had a terrestrially-based forager diet (Figure 3B). In the absence of strontium isotope analyses on each of these individuals, we cannot exclude that one or more of them were not locals, although this seems unlikely on genetic grounds in the case of UZZ71 and UZZ88, who have the same genetic makeup as the LM hunter-gatherers from G. dell’Uzzo. This may lead to the question of whether a mixed forager/fishing/farming economy may have become established at G. dell’Uzzo for up to a couple of centuries after the introduction of agropastoralism. This possibility is compatible with the zooarchaeological evidence from the site, which indicates that fishing may have been more commonly practiced in the Early Neolithic than in the Mesolithic (STAR Methods). Although rather rare, cases of HGs adopting elements of farming have been reported before in the Balkans, such as at the Iron Gates in Serbia and Romania, Malak Preslavets in Bulgaria, and very recently for the French Mediterranean coast (Bonsall et al., 2015; Forenbaher and Miracle, 2006; Gronenborn, 2017; Lipson et al., 2017b; Mathieson et al., 2018; Perlès, 2001; Rivollat et al., 2020).In addition, the few available ancestry profiles for Early Neolithic farmers in the Maghreb showed a strong population genomic continuity with the ∼15,000 yBP Iberomaurusian foragers from this region (Fregel et al., 2018; van de Loosdrecht et al., 2018). Taken together, these cases provide joint evidence of generally heightened interaction between HGs with early farmers in central and western Mediterranean regions, such as north-western Sicily (Isern et al., 2017; Mulazzani et al., 2016; Rivollat et al., 2020; Sánchez et al., 2012; van de Loosdrecht, 2021). Future research could explore the possibility that in Mediterranean coastal regions, acculturation of local foragers played a more significant role in the process of Neolithisation compared to regions along the Continental Route (van de Loosdrecht, 2021). The expansion routes into the central and western Mediterranean form an integral part of archaeological discourse on the Mediterranean Neolithic transition. Congruent with previous results for EN farmer groups in Iberia, we show that the Sicilian EN farmers also shared genetic affinity with EN groups from the Balkans, as well as groups in central Europe associated with the Continental Route, rather than North African groups from the southern Mediterranean (Figure 2B, Data S3.1, Data S5.5, Figure S10) (Haak et al., 2015; Olalde et al., 2015; Villalba-Mouco et al., 2019). It is therefore most parsimonious that the majority of early farmers in Sicily and Iberia descended from groups that expanded along a northern Mediterranean route, which share the same origin with the Continental Route in the Balkans and did not cross the Strait of Gibraltar and/or Sicily (van de Loosdrecht, 2021). Overall, our study presents genetic and dietary transitions over a 6,000 years time transect from the same archaeological site. The genomic data from individuals from Grotta del’Uzzo show evidence for at least three genetic incursions during the Late Mesolithic, Early Neolithic, and Early Bronze Age, with the most significant population shift happening during the Mesolithic-Neolithic transition. Combining genomic and isotopic evidence, we reveal that during the earliest Neolithic phase, resident HGs and incoming farmers were not only genetically interacting but also may have affected each others’ subsistence practices. It is important to note that the analysis of genomic ancestry alone may not detect acculturation. In this study and many previous reports for forager-farmer interactions, the acculturation of foragers could be detected only because of the joint analyses of genomic ancestry, stable isotope data and precise AMS radiocarbon dates together with archaeological context descriptions (Lipson et al., 2017b; Mathieson et al., 2018; Rivollat et al., 2020; van de Loosdrecht, 2021). Hence, multidisciplinary approaches form the most powerful research strategy to obtain a comprehensive understanding of the transition from foraging to sedentary farming in the Mediterranean (van de Loosdrecht, 2021). Limitations of the study An important merit of our study is to provide a view on genetic and dietary change in the early to middle Holocene at a single site in the center of the Mediterranean. Our sampling strategy succeeded in covering different phases of the Mesolithic and Neolithic, enabling us to detect key transitions in genomes and diet. However, the nature of prehistoric bone assemblages, especially when spread out across at least five millennia, makes it difficult to have at our disposal more than a few specimens for each phase. The transition from the Mesolithic to the Neolithic is a key time for our study, which has shown it to have been a short-term phase, likely lasting less than the age ranges detectable by means of radiocarbon dating. This implies that in archaeological terms, and particularly for a cave site with a complex stratigraphy and palimpsest-like deposits, we are dealing with a “needle in a haystack” scenario. Our interpretation of what may have happened around the time of contact between hunter-gatherers and early farmers thus relies mainly on genetic and isotopic data from three individuals (i.e., UZZ71, UZZ77, and UZZ81). Nevertheless, our reconstructions are not only based on the genetic and isotope data presented here but also on published isotope data (Mannino et al., 2015) as well as on a considerable amount of archaeological, zooarchaeological, and archaeobotanical research undertaken on G. dell’Uzzo (Costantini et al., 1987; Piperno et al., 1980a, Piperno et al., 1980b; Tagliacozzo, 1993; Tusa, 1985, Tusa, 1996). Another limitation that this kind of investigation has to contend with is the dearth of sites and deposits dating to the Mesolithic-Neolithic transition, which in the Mediterranean are represented by less than a handful of contexts (Biagi, 2003; Biagi and Starnini, 2016; Binder et al., 2012). Supplemental information Download .pdf (4.98 MB) |
|
|
|
Post by Admin on Jul 13, 2022 17:54:06 GMT
Genetic history of Calabrian Greeks reveals ancient events and long term isolation in the Aspromonte area of Southern Italy
Scientific Reports volume 11, Article number: 3045 (2021)
Abstract
Calabrian Greeks are an enigmatic population that have preserved and evolved a unique variety of language, Greco, survived in the isolated Aspromonte mountain area of Southern Italy. To understand their genetic ancestry and explore possible effects of geographic and cultural isolation, we genome-wide genotyped a large set of South Italian samples including both communities that still speak Greco nowadays and those that lost the use of this language earlier in time. Comparisons with modern and ancient populations highlighted ancient, long-lasting genetic links with Eastern Mediterranean and Caucasian/Near-Eastern groups as ancestral sources of Southern Italians. Our results suggest that the Aspromonte communities might be interpreted as genetically drifted remnants that departed from such ancient genetic background as a consequence of long-term isolation. Specific patterns of population structuring and higher levels of genetic drift were indeed observed in these populations, reflecting geographic isolation amplified by cultural differences in the groups that still conserve the Greco language. Isolation and drift also affected the current genetic differentiation at specific gene pathways, prompting for future genome-wide association studies aimed at exploring trait-related loci that have drifted up in frequency in these isolated groups.
Introduction
The Italian Peninsula represents a key area of investigation to explore population and demographic processes that characterized the peopling history of Europe and the Mediterranean, and to reconstruct patterns of genetic diversity at different geographical scales1. The genomic variability among present-day Italians is indeed due to the multi-layered mosaic of pre-historical and historical processes of migration and admixture that interested the continent throughout time, but also to its valuable diversity and richness in terms of ecological complexity and cultural heterogeneity. The existence of distinct environmental pressures and climate conditions were proved to have forced different patterns of local adaptations between the Northern and Southern Italy, contributing to the observed genetic structure2,3. In the same way, geographic constraints or cultural factors (e.g. language, ethnicity, socio-economic structure) were suggested to have additionally influenced human population variability along the Peninsula, being responsible for different paths of isolation and differentiation or population mobility at more fine-grained local levels4,5,6.
The distinctive genomic variability of Italy has been largely investigated by using both uniparental markers7,8,9,10, autosomal SNP-chip data2,11,12,13 and, more recently, through one of the first study on whole genome sequencing3. In this context, the analysis of geographically isolated populations or of cultural enclaves and ethno-linguistic minorities can provide a simplified observatory for exploring population relationships within and among human groups. Indeed, the condition of isolation might have helped reducing the confounding effects of admixture14,15. Furthermore, the founding event and the limited external gene flow, ultimately resulting in smaller effective population size (Ne) and increased homozygosity and linkage disequilibrium (LD), may help the study of potentially trait-associated alleles found at higher frequency in these groups, thus making population isolates key models for genome-wide association studies16,17,18,19,20. In line with this viewpoint, some of the geographic and cultural isolates today settled in the Italian territory have long attracted the attention of population genetic studies based on both uniparental and autosomal markers4,5,21,22,23,24,25,26.
Recently, a genomic survey on patterns of ancient and recent admixture in Southern Italy and the Mediterranean27 brought specific interest on the enigmatic Calabrian Greek-speaking communities residing in the Aspromonte mountain area of Bovesia, in the territory of Reggio Calabria (Southern Italy). These ethno-linguistic groups represent extant Hellenic islands in Southern Italy that still preserve a unique variety of Greek, also known as Greco or Calabrian Greek. Linguistic studies have supplied interesting information about the possible origins of these communities, historically counterpoising two main antithetic hypotheses. The first hypothesis leads back to the Medieval period and suggests that this language might derive from the descendants of Byzantines who settled Southern Italy between the fifth and eleventh centuries28,29,30,31,32. The second hypothesis instead brings to the Magna Graecia colonization of Southern Italy in the eighth century BC to trace the origin of the language. Thereafter, the Greek of Calabria would have been uninterruptedly spoken during the centuries until the present, with local developments33,34,35. More recently, reconciling scenarios tried to mitigate the initial controversy between the “Hellenic” vs. “Byzantine” dichotomy, mostly in virtue of two considerations: (i) the impact of intense Greek-Romance linguistic contacts into a dynamic model of coexistence and cohabitation between Greek-speaking Latins and Latin-speaking Greeks; and (ii) the implications of archaic lexical elements shared with peripheral Greek dialects such as those of Cyprus and the Dodecanese, to assess the importance of linguistic contributes from the Greek of different periods36,37. The new moderated hypotheses in fact consider the Greek of Magna Graecia to have been survived in diglossia with Latin during the Roman Empire and then to have been rekindled in the Byzantine era and subsequent periods. Importantly, multiple strata which repeatedly brought waves of Greek speakers onto the Calabrian coasts were suggested to have overall contributed to the Greek heritage of the region38. Historically, it is known that the Greek presence in Calabria was continuous since ancient times and that the area of Greek-influence in Southern Italy was originally more largely extended with respect to the enclaves present nowadays39. In fact, the number of Greco-speaking people is rather limited at present and today the language survives mainly in few communities residing in the Aspromonte mountain area of Reggio Calabria40,41.
Our previous characterization of Calabrian Greek communities for approximately 150,000 genome-wide SNPs with the Illumina GenoChip 2.0 DNA Ancestry chip suggested possible signs of genetic drift experienced by these groups27. In the present study, we further address issues of geographic and cultural isolation by using a higher in-depth level of analysis, which was achieved both by increasing the number of analyzed markers (720 K) and by genotyping also other communities from the same geographic area of Southern Italy (Fig. 1a). In particular, we significantly expanded the population samples from the Aspromonte mountain area by including six additional communities (Amendolea, Africo, San Lorenzo, Cardeto, Samo, San Luca) to the five already collected previously (Bova, Roghudi, Roccaforte del Greco, Gallicianò, Condofuri), therefore doubling the representativeness of the genetic structure of the area. The sampling strategy specifically covered both the communities that still preserve the Greco language as well as those from the same geographically isolated area of the Aspromonte that lost the use of this language earlier in time (Fig. 1b). This more extended sampling should therefore mirror the progressive restriction of the area of Greek-influence in the Southern Calabrian territory of Reggio Calabria. The groups from the Aspromonte mountain area were finally compared with newly-collected samples coming from a similar, but less isolated geographical context, which encompasses four villages from the Calabrian province of Catanzaro (Girifalco, Jacurso, Pentone, Tiriolo), as well as with ‘open’ (i.e. not isolated) Southern Italian groups from Castrovillari (Northern Calabria, Southern Italy) and Benevento (Campania, Southern Italy) (Fig. 1a). In this context, the aim of this study is to investigate the past population events and the local demographic factors that significantly contributed to the current genetic differentiation of Calabrian Greeks. In particular, by comparing their allelic architecture to the more general Southern Italian population we looked for the effect of geographic and cultural isolation on the detected genetic structure.
|
|
|
|
Post by Admin on Jul 13, 2022 20:56:12 GMT
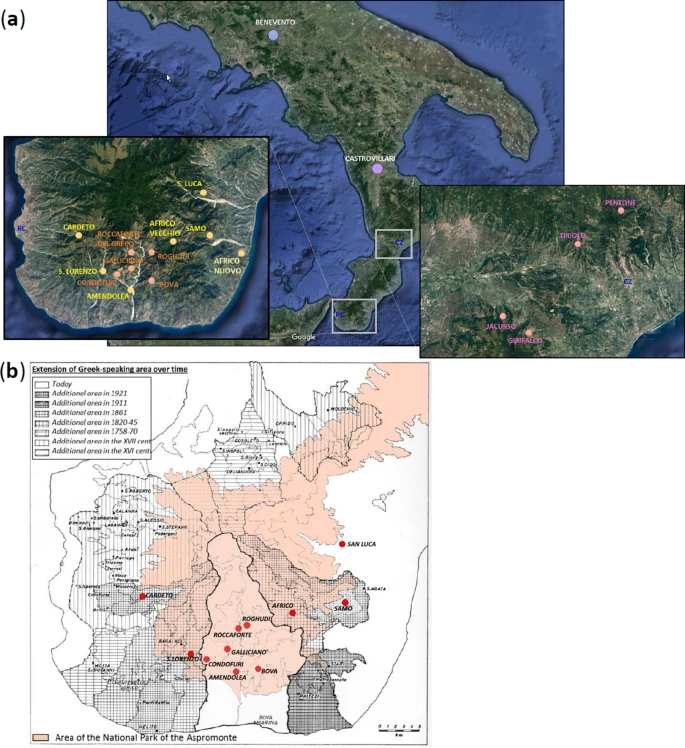 Figure 1 (a) Sampling map showing the approximate geographic location of analyzed populations. Sampling points are color-coded according to the province of origin: Benevento (blue); Castrovillari (purple); Catanzaro (magenta); previously collected samples from Reggio Calabria (orange); newly collected samples from Reggio Calabria (gold). The two enlarged boxes detail the sampling locations of villages in the province of Reggio Calabria (left) and in the province of Catanzaro (right), respectively. (b) Historical map showing the approximate extension of the National Park of the Aspromonte mountain area (in pink) as well as the range of the Greek-speaking area at different time periods as reported in the legend at the top-left. Geographical map has been generated with the package RgoogleMaps [v. 1.4.1]92 (URL: www.jstatsoft.org/v63/i04/) of the software R [v. 3.2.4] (https://www.r-project.org/). |
|
|
|
Post by Admin on Jul 14, 2022 16:14:54 GMT
Results Population structure of Southern Italy within the Euro-Mediterranean genetic landscape In order to set the observed genetic variability into a wider context, a PCA was firstly performed by comparing our newly analyzed Southern Italian populations to Mediterranean and European groups extracted from the HGDP (Suppl. Table S1). The plot of the first two principal components (Fig. 2a) recapitulates well-known geographic patterns of genetic variation commonly observed in the Euro-Mediterranean area42,43. In fact, the PC1, extending from the Levantine groups of Palestinians and Druze to Russians, Orcadians and French, identifies a South-East to North-West axis of genetic structuring. On the other hand, PC2 emphasizes the renowned outlying position of Sardinia within the European genetic landscape44,45. Populations from the Italian Peninsula reflect the known latitudinal cline of genetic differentiation between North and South Italy2,3, with the former closer to Western Europe and the latter projecting towards the Near East. Interestingly, populations from the Aspromonte area depart from the other Southern Italian groups when PC4 is considered (Fig. 2b). Accordingly, ADMIXTURE analysis identifies the main European-, Near Eastern- and Sardinian-like genetic ancestries, to which an Aspromonte-specific component is added for higher values of K (Suppl. Figure S1). Figure 2 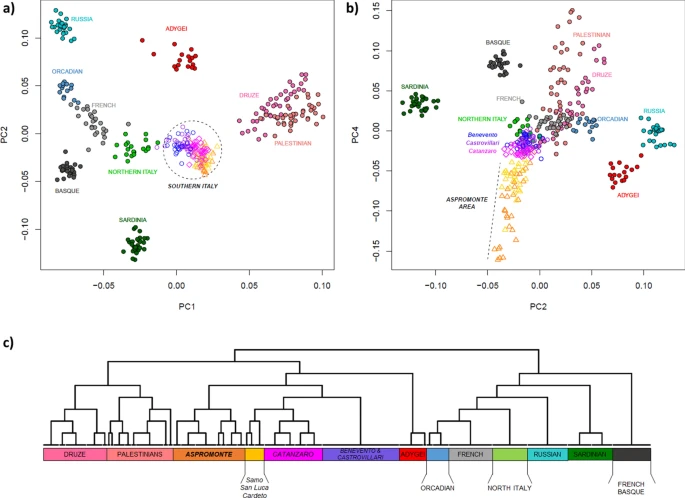 Principal component analysis and FineSTRUCTURE clustering analysis performed on the extended comparison dataset of modern populations. Scatterplots of the (a) first vs. second and (b) second vs. fourth PCs are reported on the top of the plot. Individuals are color-coded based on their geographic location. Newly analyzed populations from Southern Italy and, more specifically, those from the Aspromonte mountain area are labelled in the first and second plot respectively. (c) FineSTRUCTURE hierarchical clustering dendrogram calculated between the pairs of modern individuals included in the extended comparison dataset. The 13 detected clusters, here highlighted with different colors, are highly concordant with population labels. For the detailed annotation of individuals inside of each cluster see Supplementary Figure S3. F3-statistic was then used to formally test each Southern Italian group as target of admixture using all the other comparison population pairs as putative parental sources. Statistically significant results (i.e. Z-scores < −3) for a mixture between Sardinian and Caucasian or between Near-Eastern and continental European sources were obtained for the populations of Benevento, Castrovillari and Catanzaro (Suppl. Table S2). Consistently with these results, the maximum likelihood tree reconstructed with Treemix (Suppl. Figure S2) locates all the Southern Italian groups in an intermediate position between Caucasian and Near Eastern populations on one hand, and a continental European cluster encompassing Orcadian, Russian, French, Basque and North Italian populations together with Sardinians in a rooted position, on the other hand. When allowing for admixture, TreeMix optimized the fit of the data to the tree by adding two migration edges between populations, specifically between Russians and Adygei, and between Sardinian and French_Basque. Interestingly, adding further admixture events (from m = 3 through m = 6) increased the rate of explained variation in the data by the tree models, revealing migration edges from Caucasus (Adygei) or from a Caucasian/Near-Eastern root to the Southern Italian populations of Benevento, Castrovillari and Catanzaro. This happens each time they were included within the continental European group instead of splitting out before it (Suppl. Figure S2), thus providing further support for genetic links between Southern Italy and the Near-East/Caucasus as consistently observed by f3-admixture tests. In this context, it is noteworthy that population samples from the Aspromonte area do not show evidence of gene flow from any other group (Suppl. Table S2) and in the Treemix phylogeny appear instead located in a basal position with respect to all the other Southern Italian populations (Suppl. Figure S2). At the same time, they also show longer branch length, thus signaling possible evidence of more ancient isolation and drift effects. To empirically evaluate finer-scale patterns of structuring, we used the haplotype-based approach implemented in CHROMOPAINTER/FineSTRUCTURE to define clusters of genetically homogeneous individuals. Overall, the clusters recognized by FineSTRUCTURE (Fig. 2c) largely match with local population groups and summarize the patterns of genetic relationships consistently observed in genotype-based analyses. In fact, populations from Southern Italy globally form a clade that is related to the Caucasian Adygei group and, at a more basal level, to the Near Eastern clusters formed by Palestinians and Druze, respectively. With a more specific focus, it is worth noting how FineSTRUCTURE virtually reconnects almost all the populations from the Aspromonte mountain area to a specific cluster, which splits up as a separated group with respect to all the other Southern Italian populations (Fig. 2c, Suppl. Figure S3). Within this “Aspromonte” (ASPR) cluster, the Greco-speaking communities from Roghudi, Roccaforte del Greco, Condofuri and Gallicianò form a further sub-group that exhibits more remarkable signals of genetic drift (Suppl. Figure S4). On the other hand, samples from Cardeto, Samo and San Luca fall outside from this “Aspromonte” (ASPR) cluster and appear instead grouped to the other Southern Italian populations, namely to the cluster of villages from the province of Catanzaro (CZ) and then to a cluster indifferently encompassing all the individuals from both Benevento and Castrovillari (BN + CS) (Fig. 2c, Suppl. Figure S3). |
|

