

Post by Admin on Aug 29, 2013 7:00:58 GMT
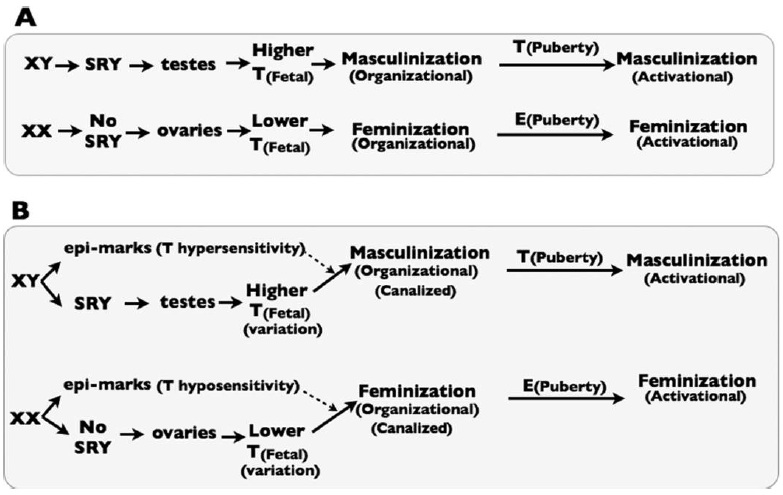
Figure 1. The Sexual Dimorphism Signaling Pathway
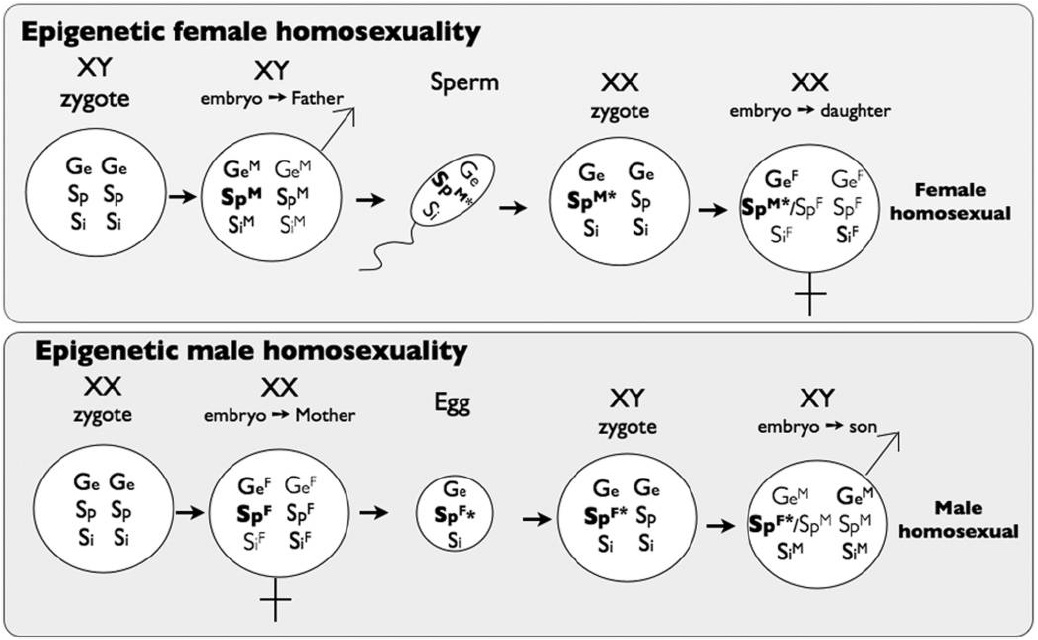
Figure 2. SA-Epi-Marks and Homosexuality
In their paper, "Homosexuality as a Consequence of Epigenetically Canalized Sexual Development", Rice et al. hypothesize that heritable epigenetic markers play a role leading to homosexuality (Rice et al., 2012). They build off of the prenatal androgen paradigm, which attributes sexual development to fetal testosterone concentrations, and add the ammendment that specific epi-marks affect fetal testosterone sensitivity (Figure 1). Usually, these epi-marks are not heritable. However, occasionally they do get passed down to the next generation. The Rice et al. model predicts that testosterone sensitizing epi-marks originate from the father, while testosterone desensitizing epi-marks originate from the mother. With increased testosterone sensitivity, an XX fetus goes through masculinization (the degree depending on multiple epi-marks and other factors). Similarly, with decreased testosterone sensitivity, an XY fetus goes through feminization to some extent. Depending on the degree of feminization of masculinization, homosexuality can theoretically result (Figure 2).
It is well established that fetal androgen signaling strongly influences sexual development. We show that an unappreciated feature of this process is reduced androgen sensitivity in XX fetuses and enhanced sensitivity in XY fetuses, and that this difference is most feasibly caused by numerous sex-specific epigenetic modifications (“epi-marks”) originating in embryonic stem cells. These epi-marks buffer XX fetuses from masculinization due to excess fetal androgen exposure and similarly buffer XY fetuses from androgen underexposure. Extant data indicates that individual epi-marks influence some but not other sexually dimorphic traits, vary in strength across individuals, and are produced during ontogeny and erased between generations. Those that escape erasure will steer development of the sexual phenotypes they influence in a gonad-discordant direction in opposite sex offspring, mosaically feminizing XY offspring and masculinizing XX offspring.
Such sex-specific epi-marks are sexually antagonistic (SA-epi-marks) because they canalize sexual development in the parent that produced them, but contribute to gonad-trait discordances in opposite-sex offspring when unerased. In this model, homosexuality occurs when stronger-than-average SA-epi-marks (influencing sexual preference) from an opposite-sex parent escape erasure and are then paired with a weaker-thanaverage de novo sex-specific epi-marks produced in opposite-sex offspring. Our model predicts that homosexuality is part of a wider phenomenon in which recently evolved androgen-influenced traits commonly display gonad-trait discordances at substantial frequency, and that the molecular feature underlying most homosexuality is not DNA polymorphism(s), but epi-marks that evolved to canalize sexual dimorphic development that sometimes carryover across generations and contribute to gonadtrait discordances in opposite-sex descendants.

