|
|
Post by Admin on Sept 18, 2014 21:51:30 GMT
Agriculture was sweeping into Europe 7,500 years ago from the Near East, bringing early farmers into contact with hunter-gatherers who had already been living in Europe for tens of thousands of years. Genetic and archaeological research in the last 10 years has revealed that almost all present-day Europeans descend from the mixing of these two ancient populations. However, researchers at Harvard Medical School in the United States and the University of Tübingen in Germany have now documented a genetic contribution from a third ancestor: Ancient North Eurasians. This group appears to have contributed DNA to present-day Europeans as well as to the people who travelled across the Bering Strait into the Americas more than 15,000 years ago. 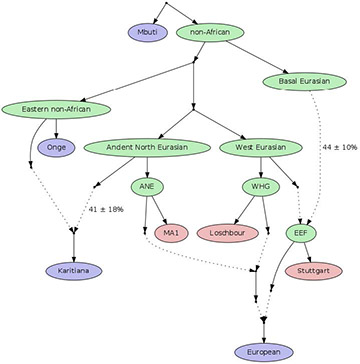 Revised flow chart of European ancestry incorporating the new data about Ancient North Eurasians (ANE), West European hunter-gatherers (WHG), early European farmers (EEF) and Basal Eurasians. Image courtesy David Reich “Prior to this paper, the models we had for European ancestry were two-way mixtures. We show that there are three groups,” said David Reich, professor of genetics at HMS and co-senior author of the study. “This also explains the recently discovered genetic connection between Europeans and Native Americans,” Reich added. “The same Ancient North Eurasian group contributed to both of them.” Basal EurasiansThe research team also discovered that ancient Near Eastern farmers and their European descendants can trace much of their ancestry to a previously unknown, even older lineage called the Basal Eurasians. The study is published in Nature. To probe the ongoing mystery of Europeans’ heritage and their relationships to the rest of the world, the international research team—including co-senior author Johannes Krause, professor of archaeo- and paleogenetics at the University of Tübingen and co-director of the new Max Planck Institute for History and the Sciences in Jena, Germany—collected and sequenced the DNA of more than 2,300 present-day people from around the world and of nine ancient humans from Sweden, Luxembourg and Germany. The ancient bones came from eight hunter-gatherers who lived about 8,000 years ago, before the arrival of farming, and one farmer from about 7,000 years ago. The researchers also incorporated into their study genetic sequences previously gathered from ancient humans of the same time period, including early farmers such as Ötzi “the Iceman.”  This skull was unearthed from a lake bed in central Sweden near the town of Motala. It formed part of the analysis that linked Northern European to indigenous hunters as well as Ancient North Eurasians Major movement of new people“There was a sharp genetic transition between the hunter-gatherers and the farmers, reflecting a major movement of new people into Europe from the Near East,” said Reich. Ancient North Eurasian DNA wasn’t found in either the hunter-gatherers or the early farmers, suggesting the Ancient North Eurasians arrived in the area later, he said. “Nearly all Europeans have ancestry from all three ancestral groups,” said Iosif Lazaridis, a research fellow in genetics in Reich’s lab and first author of the paper. “Differences between them are due to the relative proportions of ancestry. Northern Europeans have more hunter-gatherer ancestry—up to about 50 percent in Lithuanians—and Southern Europeans have more farmer ancestry.”  Scientists collected genetic data from nine ancient skeletons and 203 present-day populations living all over the world. They then isolated human DNA and sequenced the complete genomes Lazaridis added, “The Ancient North Eurasian ancestry is proportionally the smallest component everywhere in Europe, never more than 20 percent, but we find it in nearly every European group we’ve studied and also in populations from the Caucasus and Near East. A profound transformation must have taken place in West Eurasia” after farming arrived. When this research was conducted, Ancient North Eurasians were a “ghost population“—an ancient group known only through the traces it left in the DNA of present-day people. Then, in January, a separate group of archaeologists found the physical remains of two Ancient North Eurasians in Siberia. Now, said Reich, “We can study how they’re related to other populations.” A greater complexityThe team was able to go only so far in its analysis because of the limited number of ancient DNA samples. Reich thinks there could easily be more than three ancient groups who contributed to today’s European genetic profile. He and his colleagues found that the three-way model doesn’t tell the whole story for certain regions of Europe. Mediterranean groups such as the Maltese, as well as Ashkenazi Jews, had more Near East ancestry than anticipated, while far northeastern Europeans such as Finns and the Saami, as well as some northern Russians, had more East Asian ancestry in the mix. The most surprising part of the project for Reich, however, was the discovery of the Basal Eurasians. “This deep lineage of non-African ancestry branched off before all the other non-Africans branched off from one another,” he said. “Before Australian Aborigines and New Guineans and South Indians and Native Americans and other indigenous hunter-gatherers split, they split from Basal Eurasians. This reconciled some contradictory pieces of information for us.” Next, the team wants to figure out when the Ancient North Eurasians arrived in Europe and to find ancient DNA from the Basal Eurasians. “We are only starting to understand the complex genetic relationship of our ancestors,” said co-author Krause. “Only more genetic data from ancient human remains will allow us to disentangle our prehistoric past.” “There are important open questions about how the present-day people of the world got to where they are,” said Reich, who is a Howard Hughes Medical Investigator. “The traditional way geneticists study this is by analysing present-day people, but this is very hard because present-day people reflect many layers of mixture and migration. “Ancient DNA sequencing is a powerful technology that allows you to go back to the places and periods where important demographic events occurred,” he said. “It’s a great new opportunity to learn about human history.” |
|
|
|
Post by Admin on Sept 24, 2014 20:33:35 GMT
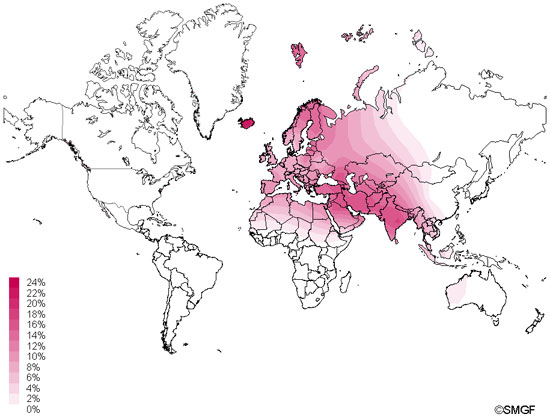 Haplogroup U is found in 15% of Indian caste and 8% of Indian tribal populations.[2] Haplogroup U is found in approximately 11% of native Europeans and is held as the oldest maternal haplogroup found in that region.[2][3][4] In a 2013 study, all but one of the ancient modern human sequences from Europe belonged to maternal haplogroup U, thus confirming previous findings that haplogroup U was the dominant type of Mitochondrial DNA (mtDNA) in Europe before the spread of agriculture into Europe.[5] Haplogroup U is subdivided into Haplogroups U1-U9. Haplogroup K is a subclade of U8.[6] The old age has led to a wide distribution of the descendant subgroups across Western Eurasia, North Africa, and South Asia. Some subclades of U have a more specific geographic range.  A Kalash girl of the Chitral district in North Pakistan Haplogroup U4 has its origin in the Upper Palaeolithic, dating to approximately 25,000 years ago and has been implicated in the expansion of modern humans into Europe occurring before the Last Glacial Maximum.[31] U4 is an ancient mitochondrial haplogroup[32] and is relatively rare in modern populations.[33] U4 is found in Europe with highest concentrations in Scandinavia and the Baltic states[34] and is also associated with the remnants of ancient European hunting-gatherers preserved in the indigenous populations of Siberia.[35][36][37] U4 is found in Nganasans the indigenous inhabitants of the Taimyr Peninsula,[13][38] in the Mansi (16.3%) an endangered people,[37] and in the Ket people (28.9%) of the Yenisey River.[37] U4 is also preserved in the Kalash people a unique tribe among the Indo-Aryan peoples of Pakistan (current population size 3,700)[39] where it attains its highest frequency of 34%.[40][41][42] Haplogroup U4 is associated with ancient European hunter-gatherers and has been found in 7,200 to 6,000-year-old remains of the Pitted Ware culture in Gotland Sweden and in 4,400 to 3,800-year-old remains from the Damsbo site of the Danish Bell-Beaker culture.[48][49][50] Remains identified as subclade U4a2 are associated with the Battle Axe culture which flourished 5,200 to 4,300 years ago in eastern and central Europe and encompassed most of continental northern Europe from the Volga River in the east to the Rhine River in the west.[51] Mitochondrial DNA recovered from 3,500 to 3,300-year-old remains at the Bredtoftegård site in Denmark associated with the Nordic Bronze Age include haplogroup U4 with 16179T in its HVR1 indicative of subclade U4c1.[50][52][53][54] 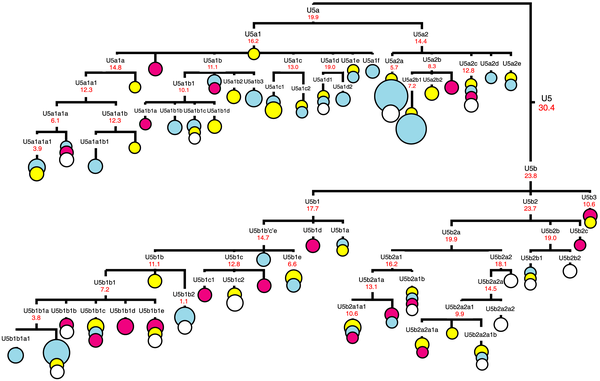 Figure 1. Complete mtDNA phylogenetic tree of haplogroup U5. It has been suggested that hg U5, or a genetically close ancestor to U5, arose among the first settlers of Europe, and subsequently spread all aver the continent along with the Aurignacian industry [1],[2]. According to archaeological data, this Upper Paleolithic industry spread rapidly across western and central Europe roughly 42 to 40 kya, and there is evidence for a slightly earlier influx in southern and central Europe [12]. It is also suggested that Upper Paleolithic occupation at archaeological sites on the Don River in the East European Plain (at Kostenki site) occurred a little earlier, around 45–42 kya [13]. However, taxonomic assignment of paleoanthropological material (teeth) from this site, as well as the relationship between the Kostenki industry and the earliest dated Upper Paleolithic remains in southern and central Europe, remains unclear [13]. A recent study of ancient DNA from the Kostenki site (~30 kya) in Russia has shown that anatomically modern humans with haplogroup U2 might have populated the East European Plain at that time [14]. Therefore, as in southern and central Europe, only the youngest of the archaeological entities (Aurignacian) in eastern Europe is associated with skeletal remains that may be assigned unequivocally to anatomically modern humans [15],[16]. It is probable, however, that the first successful expansion of modern humans inhabiting the territory of the central part of East European Plain might have occurred at least 24 kya [17]. This follows from the fact that the density of precisely dated sites increased considerably in southern Russia during the Last Glacial Maximum (roughly 24–16 kya), and in north western Russia after degradation of the glacier ice ~14 kya [18]. Therefore, the hypothesis of the eastern ‘Periglacial refugium’ postulated based on archaeological data [18],[19] is probably supported by genetic data presented here, as the coalescence times for some U5a subclusters (as well as for hg U4 [20]) in eastern Europe correspond to the post-LGM dates. However, we did not detect any mtDNA haplotypes of such antiquity that clearly identify early traces of the pre-LGM expansions in eastern Europe, originating from industries that can be traced back 40–30 ky (for instance, for the Spitsyn or Strelets Cultures assemblages). Evolutionary ages of ~20–24 ky have been found for ancestors of subhaplogroups U5b and U5b2, which most probably were originated in southern and central part of Europe, because the majority of such haplotypes were revealed in Czechs, Slovaks and Poles and in the Mediterranean populations, based on a set of population data analyzed in the present study (Figure 1, Figure S2). It is known that the Late Upper Paleolithic industries of central Europe are closely related to contemporaneous eastern European cultures [21],[22]. This probably reflects both the proximity of central Europe, and the presence of similar environments in the Carpathian Basin. The eastern Gravettian industry, which is represented in sites dated back ~29–22 kya, is found in both central and eastern Europe [22]. However, climates were milder in the Carpathian Basin and, in contrast to the central East European Plain, settlements in central Europe appear to be continuous throughout the LGM [21]. Moreover, the eastern part of central Europe - the Carpathian Basin, is considered by archaeologists and ecologists as one of the European refugia that existed during the LGM [23],[24]. A recent ancient DNA study of Stone Age hunter-gatherers from central and eastern Europe has shown that most of the samples studied (>80%) shared mtDNA haplotypes belonging to haplogroups U5 and U4, haplogroups that notably are relatively rare in central Europe today [7]. Among them, sample 5830a from Hohlenstein-Stadel (Germany), defined as U5a2a, based on the HVS I sequence 16114A-16192-16256-16294-16311, was detected. Its closest HVS I relatives in modern populations can be found among eastern Europeans (in Latvians, Russians, Tatars and Mordvins). In the present study we have completely sequenced two U5a2a mitochondrial genomes with the characteristic transition at np 16311 in Russians (Figure S1). AMS radiocarbon dating of sample 5830a (at ~7.8 ky old) allows determination of a minimum age for the subcluster U5a2a to which it belongs. Phylogenetic analysis revealed that the coalescent ages of this subcluster vary from 5.7±1.2 ky (for a complete mtDNA clock rate) to 9.3±2.6 ky (for a synonymous clock rate). Therefore, the age of sample 5830a falls within the intervals of these molecular phylogenetic dating. We should note, however, that Russian U5a2a-16311 samples share a mutation at np 9293 with a Belorussian sample, thus forming a subcluster with the age of ~2.6 ky (Figure S1). Thus it is probable that similar HVS I sequences in Stone Age individuals and modern Russian individuals may be related only within the bounds of the whole subcluster U5a2a, defined, in fact, by parallel mutations at np 16311 originating independently. In addition, another ancient HVS I haplotype, 16192-16270, belonging probably to subcluster U5b1a, was found in skeletons dated to ~5.6 ky old, from Lithuania [7]. We have found that the age of ancient mtDNAs does not exceed the coalescence time estimates of modern U5b1a sequences, falling within the range of 9.3±3.5 ky (for a complete mtDNA clock rate) to 6.8±4.8 ky (for a synonymous clock rate) (Figure S2). In general, it seems that the combining of phylogenetic and archaeological approaches will be useful for cross-checking data and improvement of human mitochondrial molecular clock estimates, as new information about modern and ancient mitochondrial genomes becomes available [3]. Genetic data obtained in this study allows the suggestion that during the LGM period, central European territories probably represented the area of intermingling between human flow from refugial zones in the Balkans, the Mediterranean coastline and the Pyrenees, as U5a and U5b gene flows occurred from there. Based on dating analysis of the U5 subclusters, it seems very likely that, despite the archaeological evidence testifying to the presence of humans in eastern Europe during the Ice Age, this part of Eurasia might have only been re-populated by modern humans at the end of the LGM, i.e. later than central Europe. In addition, U5b gene flow from central to eastern Europe become much more intense after the LGM. In general, we believe that molecular genetic data, in addition to archaeological and fossil evidence, are of significant use for resolving key questions regarding the interaction of human communities and climate. Malyarchuk, Boris, et al. " The peopling of Europe from the mitochondrial haplogroup U5 perspective." PLoS One 5.4 (2010): e10285. |
|
|
|
Post by Admin on Sept 25, 2014 20:51:03 GMT
A multinational team analysed stone tools recovered during a recent re-excavation of the find site of the Venus of Willendorf in Austria. The team, led by Philip Nigst (University of Cambridge and Max Planck Institute for Evolutionary Anthropology) and Bence Viola (Max Planck Institute for Evolutionary Anthropology) identified the stone tools as belonging to the Aurignacian culture, generally accepted as indicative of modern human presence. Chronostratigraphic information suggests the tools date to around 43,500 years ago, pre-dating other known Aurignacian artefacts. Based on the type of soil and its mollusk assemblage, climatic conditions during that time were likely cool, with a steppe-like environment and some conifer trees along river valleys. Shared the regionThe date of the artefacts represents the oldest well-documented occurrence of behaviourally modern humans in Europe and suggests contemporaneity with Neanderthals in other parts of Europe, showing that behaviourally modern humans and Neanderthals shared this region longer than previously thought. Additionally, the results suggest that the early modern human settlers, who may have come from the warmer environments in southern Europe, were well-adapted to a variety of climates, according to the authors. Modern humans dispersed into Europe and replaced Neanderthals at least 40 thousand years ago, but potentially much earlier. “The problem is that we have practically no human remains associated with the early Upper Palaeolithic in Europe, so we have to use archaeological proxies to figure out when the first modern humans appeared. We have some clearly modern human remains associated with the Aurignacian culture, so we think it is a good indicator for modern human presence,” explains Bence Viola.  Aurignacian populations of modern humans spread mainly along the Danube valley, whereas Proto-Aurignacian populations spread along the Mediterranean coast. “At Willendorf, we could date the early Aurignacian to 43,500 years, quite a bit earlier than elsewhere and overlapping with directly dated Neanderthal remains,“ says Philip Nigst. Neanderthals’ capabilities are still hotly debated. Some argue that before modern humans replaced them, Neanderthals showed cultural capabilities similar to those of modern humans, while others make a case that these similarities only appear once Neanderthals came in contact with modern humans.  Contact with modern humans Contact with modern humans“The new data from Willendorf clearly shows that modern humans were present in what is now Austria while Neanderthals still occupied other regions of Europe suggesting that the two species met, and may have exchanged mates and ideas”, explains Philip Nigst. “This means the changes in the material culture of some of the last Neanderthal groups are most probably related to direct or indirect contact with modern humans,” says Jean-Jacques Hublin, director of the Department of Human Evolution at the Max Planck Institute for Evolutionary Anthropology. Steppe-like climateThe stone tools were discovered in a sequence of sediments that were deposited during different colder and warmer phases within the last glacial. Based on the type of soil and its mollusk assemblage, climatic conditions during the time of modern human occupation were likely cool, with a steppe-like climate and some conifer trees along river valleys. 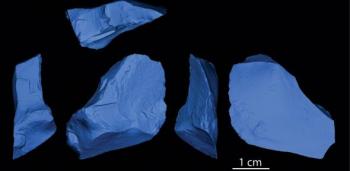 Radiocarbon dating with stratigraphy show the stone tools - including small "bladelets" - date from 43,500 years ago. “Mollusks are great for environmental reconstruction because they are so sensitive to changes in temperature and moisture – meaning that the species you find vary with every little change in climate,” explains Bence Viola. “What is particularly interesting is that the Aurignacian at Willendorf occurs in a relatively cold period, which shows that these earliest settlers were already adapted to different climates requiring different subsistence strategies,” says Philip Nigst. |
|
|
|
Post by Admin on Oct 11, 2014 20:45:56 GMT
In Western Europe, Early and Middle Pleistocene sites that have produced human fossils generally reflect an earlier settlement of the Mediterranean region compared to northern Europe [1]–[11], despite the number of both recent and previous finds (Figure 1) coming from Germany [12]–[14] or England [15], [16]. Moreover, human fossils from the loessic plains or valleys of northern France remain extremely rare, limited to finds from Biache-Saint-Vaast [17]–[20]. Here, we report new human fossils from Tourville-la-Rivière (Seine-Maritime, France) that fill both geographic and chronological gaps in our understanding of this important period in European prehistory. Three left upper limb diaphyseal sections, most likely belonging to a single individual, were found in September 2010 during rescue excavations of this Middle Pleistocene site. This new find provides insight concerning the relationship of the Tourville remains to other Middle Pleistocene human fossils [12], [21]–[24]. We have applied U-series analyses on the human bones and combined US-ESR dating on faunal teeth to refine the chronology of Tourville-la-Rivière. 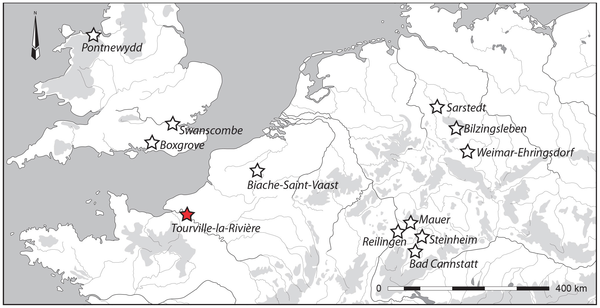 Figure 1. Location of the open-area site of Tourville-la-Rivière and other Northwest European (north to 45°N and west to 16°E) contexts, contemporaries of lower and middle Pleistocene (MIS-10-6), that have yielded human remains. Figure 1. Location of the open-area site of Tourville-la-Rivière and other Northwest European (north to 45°N and west to 16°E) contexts, contemporaries of lower and middle Pleistocene (MIS-10-6), that have yielded human remains.The open-air site of Tourville-la-Rivière was discovered in 1967 in a Seine Valley gravel quarry (Figure 2A) that has been assiduously monitored by archaeologists, with several excavations having produced Early and Middle Palaeolithic faunal and lithic assemblages [25]–[33]. The site's substantial archaeological sequence lies on the lower terrace of the Seine River, abutting a chalky Cretaceous cliff (Figure 2B), which protected this >30 m thick geological formation. 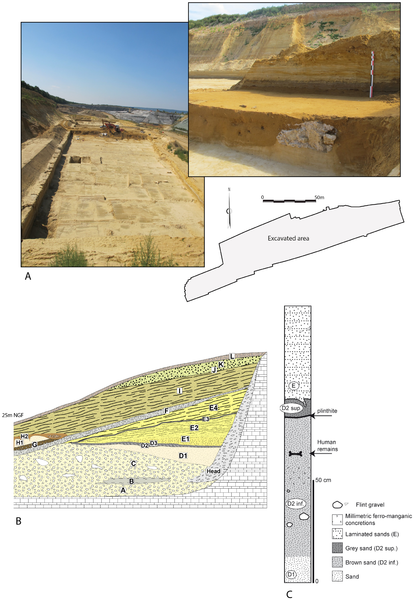 Figure 2. The Tourville-la-Rivière site. Figure 2. The Tourville-la-Rivière site.
A: general view of the site during excavation; B: general stratigraphy of Tourville-la-Rivière (after [36], modified); C: stratigraphy of the excavated area.This stratigraphic sequence, in the heart of a large meander of the Seine, comprises alluvial, estuarine, and continental sediments deposited from MIS 10 to 2 [34]–[38]. The majority of these deposits were lain down during the Saalian (MIS 8 to 6, or ~300 ka to 130 ka) [39]–[41], making Tourville a reference sequence for Middle Pleistocene environmental change in northwestern Europe. The lowermost deposits are composed of coarse periglacial gravels and sands (layer C), overlain by fine-grained alluvial sediments (sands and silts), which are sub-divided into three layers (D1, D2 and D3). The upper-part of the sequence contains laminated sands (layer E) topped by periglacial deposits (layers F to K) composed of slope deposits and aeolian sands. Based on malacological analyses [42], [43], the white sands comprising sedimentary sub-unit D1 accumulated during full interglacial conditions associated with the development of forest biomes. The top of D1 and D2 are dominated by snail species preferring more open habitats, suggesting a transition to a cold climatic period. Finally, D3 is characterised by species typical of cold and humid phases, clearly indicative of an Early Glacial phase. The role of the Seine River in the deposition and remobilisation of the Tourville fauna is unquestionable. Although it is clear that the faunal assemblage derives from multiple agents (natural processes, large carnivores, humans), it has been impossible to untangle their respective contributions. Nevertheless, bone splinters and green-stick fractures characteristic of marrow processing demonstrate the anthropic nature of the faunal material in certain excavation zones. As with the faunal remains, the lithic artefacts are spread across the excavated area, separate from a small 9 m2 zone that represents a knapping concentration (Figure S3 in File S1). All of the raw material employed is local Senonian flint collected from the chalk cliff or local alluvium. The assemblage is composed of small pieces (chips, debris), core management flakes (cortical flakes or éclats débordants), rare non-Levallois cores, retouched tools (notches, becs, scrapers) or finished products, notably Levallois blanks and non-Levallois blades. The total absence of Levallois cores and the recovery of isolated Levallois products (elongated bi- and unipolar flakes) provide evidence for the substantial fragmentation of the reduction sequence and the importation of Levallois products to this zone [44] (Text S1 in File S1). Several refit sequences demonstrate that cores and especially the largest unbroken products were transported away from the knapping zone (Figure S4 in File S1). Additional elements present in this zone can be connected to a slightly different Rocourt-type technology [45]–[49], which produced laminar flakes as well as blades (Text S2 and Figure S5 in File S1). Despite certain conceptual similarities with Levallois blade production, this Rocourt system exploits the core's center rather than surface. Although the objective is the production of elongated flakes and blades, this method also differs from Upper Palaeolithic-like Mousterian blade technology well-known during the Early Weichselian (MIS 5d-5a or 110 ka–70 ka) of northern Europe [49]–[55]. In this same region, Rocourt-type debitage is known from sites coeval with the beginning of the Weichselian (MIS 5d-5a) [56], [57]. The presence of this debitage method at Tourville thus provides yet another early example of this technology. The small number of artefacts combined with the limited size of the single knapping concentration suggest rather ephemeral human occupations spread over a fairly substantial activity area [44], [58]. Elongated Levallois flakes and non-Levallois blades were likely designed for use in butchery or carcass processing, a probability reinforced by a preliminary functional analysis concerning a sample of these artefact types produced outside the excavated area [59] (Text S3 and Figure S6 in File S1). The Tourville human remains were discovered on September 10, 2010 (Figure 3) by A. Cottard and A. Thomann. Minor damage sustained during excavations and some taphonomic alterations are evident, and the postero-lateral portion of the lower third of the diaphysis is missing. The three shaft sections were oriented in approximately the same direction, which is common for elongated elements deposited in fluvial contexts or water lain deposits [62]–[64]. The combination of archaeothanatological inferences (the order in which the articulations dislocate) [65], the susceptibility of the bones to fluvial displacement, and an anatomical study (see below), suggest the most parsimonious scenario being the fluvial transport of the complete upper limb (with or without the hand), with subsequent minor post-depositional displacement and more dramatic damage affecting the arm and forearm. If the hand had been transported along with the three shaft fragments, the presence of faunal remains in anatomical position and their differential preservation (see above and Figure S1 in File S1) complicates an explanation for its absence. 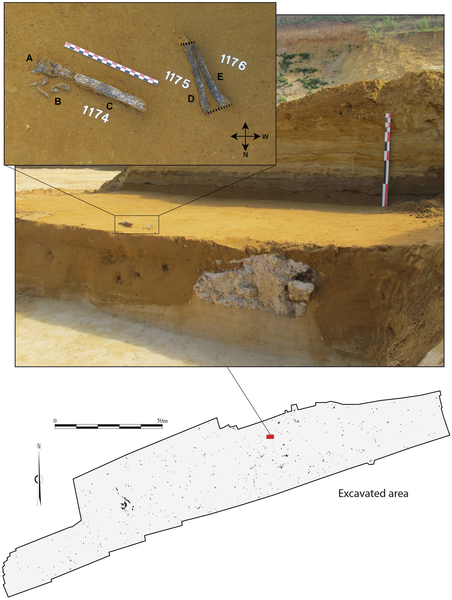 Figure 3. The Tourville human remains in situ. Figure 3. The Tourville human remains in situ.
The posterior and medial surfaces were the first to be made visible for the radial (# 1175) and ulnar (# 1176) diaphyses, respectively, while the postero-medial surface of the humeral diaphysis (#1174) and posterior surface of the distal extremity were the first to be exposed. A: distal extremity of the humerus, posterior face; B: fragments of the distal portion of the humeral diaphysis. Several elements have since been refitted to the diaphysis (see Figure 4); C: the humeral diaphysis, medial to posteromedial face, proximal extremity to the north-west; D: radius, posterior face, proximal extremity to the north; E: ulna, medial face, proximal extremity to the north. Dotted lines indicate the alignment of the broken part of the distal and proximal extremities of the ulna and radius.The three incomplete bones belong to a left humerus, radius, and ulna that were partially crushed but have been restored and reconstructed (Figure 4, Text S5 in File S1). Their external cortical surface is heavily altered, stained dark grey to black and interspersed with small white patches, a coloration affecting the entire thickness of the cortical bone. This most likely results from depositional conditions tied to a hydromorphic sedimentary regime (standing water [62]–[64]). Small, rounded cracks are also visible, and their occasional star-like organisation may be a product of root etching [66]. Given their dimensions, these bones most likely belong to an adult or an older adolescent. Data concerning the comparative samples used in the morphometric analysis are described in File S1 (Text S6, Table S3 in File S1).  Figure 4. The Tourville left upper limb remains. Figure 4. The Tourville left upper limb remains.
Top: humerus; bottom left: ulna; bottom right: radius. For all the bones: A: anterior view; M: medial view; P: posterior view; L: lateral view.Taxonomic Attribution of the Tourville Human Fossil
Discussions regarding the Middle Pleistocene emergence of the Neandertals in Europe [12], [23], [90], [91] are primarily focused on cranial and dental autapomorphies, since few of their post-cranial features appear to be derived relative to earlier Pleistocene humans. This is particularly problematic for northern Europe, where the lack of comparative remains has limited the taxonomic attribution of post-cranial remains to being described simply as non-modern Homo (e.g. the Boxgrove tibia [16]). Although the Tourville human remains conform to the general Neandertal morphological pattern, they are insufficient by themselves to provide a secure taxonomic attribution. Yet, given the presence of uniquely derived Neandertal traits on the contemporaneous Biache-Saint-Vaast specimens [19], [92], it is therefore appropriate to place the Tourville fossil in the Neandertal lineage. Behavioural InterpretationAn unusual skeletal morphology, hitherto unknown for a Pleistocene fossil, is evident on the Tourville humerus, the abnormal bone formation at the deltoid tuberosity. An hypertrophied deltoid tuberosity is evident on a (probably Neandertal) right humerus from Khvalynsk [93] and on the left humerus of the Saint-Césaire 1 Neandertal [94]. Neither of these humeri, however, exhibits the kind of entheseal change evident on the Tourville humerus. Yet, at least one humerus (humerus III) from the Sima de los Huesos may have a similar crest. Various causes can explain the crest on the Tourville humerus. Despite the multifactorial aetiology of entheseal changes in modern populations [95], we consider the simplest explanation for the altered muscular attachment to be biomechanical, with the enthesophyte at the summit of the humeral crest resulting from a single, more ‘violent’ trauma. The overall crest formation most likely results from repetitive micro- and or macro-traumas connected to the synergistic stabilization the arm associated with abduction and extension. Although the exact motion responsible for this entheseal change is difficult to determine, actions connected to throwing seem plausible [71], especially given the need for glenohumeral stability in spear throwing [96], as has been suggested for several Middle Palaeolithic contexts [cf.], [ 97]–[99]. Finally, there is a growing body of evidence for Middle [100]–[106] and Late [87], [106]–[115] Pleistocene non-modern human serious skeletal developmental variations, or minor ones, and associated survivorship. The Tourville humeral abnormality provides an additional case of a specific skeletal degeneration, which, in this case, is probably related to a specific activity or set of activities. ConclusionRescue excavations at the site of Tourville-La-Riviere produced substantial lithic and faunal material as well as a left humerus, ulna and radius belonging to the same individual and attributable to the Neandertal lineage. The site preserves a series of ephemeral but specialised MIS 7 occupations probably focused on butchery activities. The extensively excavated area (>2.5 acres) provides a window on a large part of the late Middle Pleistocene river valley, where humans transported stone tools between areas, discarding particular implements either where new ones were produced and then exported for later use or in locations where they were briefly used. This techno-economic data portrays a significant fragmentation of the reduction sequences [58] and a high mobility of the artefacts within the local environment of the Seine River valley. While it is impossible to trace the taphonomic history of the human remains, their spatial organisation and anatomic proximity are similar to some of the faunal remains. In the absence of evidence for human or carnivore intervention, the most straightforward explanation for the presence of a human left arm at Tourville is its introduction to the site by fluvial transport. The morphological and metrical comparisons demonstrate this fossil to fall within the variability of the Neandertal lineage. Moreover, the Tourville humerus represents the first case of an unusual crest at the attachment site of the posterior deltoid muscle for a Pleistocene fossil. Finally, the Tourville fossils are not only the oldest found in France during a rescue excavation, but also provide new material to what remains an extremely limited fossil sample from northwestern Europe, particularly in terms of post-cranial elements. Moreover, the trauma evident on the Tourville humerus may shed light on Neandertal behaviour. One possible explanation for the entheseal remodelling of the posterior deltoid muscle insertion is the habitual loading and torsional strain of the shoulder, possibly connected to repetitive movement. While interesting, this entheseal change probably had little bearing on the survival of the individual. The possible origins of this trauma may pose interesting questions about behavioural patterns among earlier Middle Palaeolithic humans. Faivre J-P, Maureille B, Bayle P, Crevecoeur I, Duval M, et al. (2014) Middle Pleistocene Human Remains from Tourville-la-Rivière (Normandy, France) and Their Archaeological Context. PLoS ONE 9(10): e104111. doi:10.1371/journal.pone.0104111 |
|
|
|
Post by Admin on Oct 20, 2014 1:17:59 GMT
Haplogroup E1b1b (Y-DNA)Haplogroup E1b1b (formerly E3b) represents the last major migration out of Africa into Europe. It is believed to have first appeared in the Horn of Africa or southern Africa approximately 26,000 years ago and dispersed to the Middle East during the Upper Paleolithic and Mesolithic periods. On the European continent it has the highest concentration in north-west Greece, Albania and Kosovo, then fading around the Balkans, the rest of Greece and Western Turkey. Outside Europe, it is also found in most of the Middle East, northern and eastern Africa, especially in Morocco, Lybia, Egypt Yemen, Somalia, Ethiopia and South Africa.  E1b1b1a (or E-M78, formerly E3b1a) is the most common variety of haplogroup E among Europeans and Near Easterners. E-M78 is thought to have migrated out of Egypt in the early Neolithic to colonise the Levant, Anatolia and Greece, where it mixed with the J2 inhabitants. Near-Eastern farmers settled in northern Greece circa 8,500 years ago, launching the Thessalian Neolithic (6500-2500 BCE). Looking at a map of E1b1b1a, one would almost think that this lineage came straight from Egypt to Greece, or at least from the southern Levant to Greece. The lower incidence of E1b1b1a in Syria and Anatolia is almost certainly due to the competition from the other major Neolithic haplogroups : G2 and J2.  E-M78 is divided into 4 main branches : E1b1b1a1 (E-V12), E1b1b1a2 (E-V13), E1b1b1a3 (E-V22) and E1b1b1a4 (E-V65), each further subdivided in "a" and "b" subclades. E-V13 is one of the major markers of the Neolithic diffusion of farming from the Levant. Like all the other subclades of E1b1b1a, E-V13 originated in North-East Africa around the end of the last Ice Age. Its frequency is now far higher in Greece, South Italy and the Balkans than anywhere else due to a founder effect, i.e. the migration of a small group of settlers carrying mostly this lineage (but also a small amount of other North-East African lineages, notably E-M123 and T). Archeological evidence shows that the region of Thessaly, in northern Greece, was the starting point (circa 6,000 BCE) for the diffusion of agriculture through the Balkans and the Danube basin, as far as northern France to the west and Russia to the east. The modern distribution of E-V13 hints at a strong correlation with the Neolithic and Chalcolithic cultures of Old Europe, such as the Vinča, Boian, Maritsa and Karanovo, cultures. Owing to this early introduction to Europe, E-V13 is now by far the most common E subclade found among Europeans. E-V13 is also associated with the ancient Greek expansion and colonisation. Outside of the Balkans and Central Europe, it is particularly common in Southern Italy, Cyprus and Southern France, all part of the Classcical ancient Greek world. E-V22 is the predominant subclade in the Levant and is therefore associated with the Phoenicians and Jews, in addition to the spread of agriculture. The Phoenicians could have spread E-V22 to Sicily, Sardinia, southern Spain and the Maghreb, and the Jews to Greece and mainland Italy and Spain. However, the Mediterranean route for the diffusion of agriculture (see map below) went through the exact same regions. It is therefore impossible to know at present which of the two periods (Neolithic or Classical Antiquity) played the stronger role in the spread of V22 around the Mediterranean.  The Red Sea admixture peaks in Ethiopia and Somalia, the region of origin of Y-DNA haplogroup E1b1b, to which its distribution is closely linked, except in northwestern Europe. E-V12 is the most common subclade of M78 in Egypt. Its low presence around Greece and Anatolia indicates that it probably already existed when E moved there in the early Neolithic. E-V65 is found in North Africa, with a maximum frequency in Lybia, then Morocco. It is also likely to have originated in Egypt. In Europe it is found at low frequencies in Greece and Sicily, but interestingly makes up one fourth of Sardinian E. It could be due to immigration from the Phoenician colonies in the Maghreb to Sardinia (the Sardinian haplogroup I2a1 is also present at low frequencies along the coast of Algeria and Tunisia, confirming exchanges of population between the two regions, maybe when both were Phoenician colonies). E1b1b1b (E-M81, formerly E3b1b) is characteristic of the Berbers of North-West Africa. In some parts of Morocco E1b1b1b can peak at 80% of the population. This sub-hapolgroup is also found in Iberia, Italy and southern France, with the highest concentrations in southern Portugal (12%) and decreasing as we move north.  E1b1b1c (E-M123) and its main branch E1b1b1c1 (E-M34) is also associated with the diffusion of agriculture. This haplogroup peaks in the southern Levant (10-12% in Palestine and Lebanon), from where it expands in all directions over the Middle East, North Africa, South Asia and South-East Europe. The distribution of E-M123 matches almost exactly the expansion of farming (see map below) during the Neolithic period. E-M123 seems to go hand in hand with haplogroup G and J2, with the difference that G and J2 both reach their maximum frequencies around the Caucasus and Anatolia, where cattle, pigs and goats where first domesticated. Inside Europe, E-M123 follows more or less the distribution of E-V13, with the highest frequency (1 to 5%) observed in Greece, South Italy, the Balkans and the Danube basin, then fading towards Germany, Poland, Ukraine and Russia, where its frequency is under 1%.
|
|

