|
|
Post by Admin on Sept 3, 2020 22:20:41 GMT
7.3 On the Keilmessergruppen/M.M.O. ecological and cultural implication This study, though considering only G-Complex backed tools, provides data supporting the main interpretation by Richter, for which Mousterian and Micoquian technological traditions are actually deeply interrelated, according to his M.M.O. definition [9]. The Keilmessergruppen/M.M.O. could be framed within the Mousterian variability of Late Middle Paleolithic, since typological differences or the presence of bifacial technologies don’t represent a sufficient cultural discriminating factor. Clear examples, in this sense, are Sesselfelsgrotte G-Complexes, along with the other main Keilmessergruppe/M.M.O. assemblages, where the technological and typological Mousterian substrate is always present or dominant, except in a few short-term and specialized contexts [15,47,52]. Within Keilmessergruppen/M.M.O., however, the relation between the so-called Mousterian “common background” and the “Micoquian option”, is difficult to interpret. The sites length occupation hypothesis isn’t always adaptable, since Micoquian option appears to be related to frequent/high mobility strategies, thus emerging in contexts characterized by marked ecological constraints that could match with both long-term and short-term occupation. This dichotomy cannot therefore be explained only with a tool’s reduction factor directly related to the type of occupation [127]. There is indeed a deeper element of ecological adaptation and predetermination of behavioral strategies, within which a different concept of technical object fits. The Keilmesser, that in its variety of forms and concepts represents the main technical element of Keilmessergruppen/M.M.O., is indeed a precise ecological guide-fossil; it is the versatile, multi-purpose and strategical tool adapted to constrained situations. This fossil and the entire “option” emerge with nomadic seasonal occupations with cold or even harsh climates, characterized by the “mammoth steppe” environment, with the common presence also of reindeer, horse and rhinoceros [26,47]. These constrained landscapes represent a common denominator of Keilmessergruppen/M.M.O. human occupations, while those more properly Mousterian (without the “Micoquian” option) can be associated with broader periods and areas, often including milder environments. This is true in a global Pan-European context, but is noticeable also within the same sites: in this sense, an example could be the interstratification of open-air sites in Königsaue. Königsaue-A and Königsaue-C, characterized by Levallois method next to Keilmesser and bifacial reduction, are interframed by Königsaue-B, where unifacial tools and core-reduction are exclusive. Here, raw materials management and other behavioral factors don’t record marked differences, noticeable mainly in the frequency of bifacial and unifacial technologies [15]. Whether the stratigraphy represents the whole last glacial cycle [33] or climatic oscillations within early MIS 3 [47], the layer characterized by high rate of bifacial tools (K-A) record more harsh conditions, while layers where bifacial reduction is absent (K-B) or present but rare (K-C) takes place during a milder interstadial with forestal covering(Königsaue-A) or presence (Königsaue-C) [111]. If we take as example an interregional context, in the comparison between Polish, Moravian and Slovakian sites by Burdukiewicz [105], better ecological conditions characterized the regions south of the Carpathians, where the backed bifaces complexes are rarer even though the Middle Paleolithic sites are numerous. An ecological preference was therefore recognized, with Neanderthal groups going south mainly in colder periods. In Central-Eastern Europe the Ciemna stratigraphical sequence is particularly important, where the beginning of Keilmessergruppe/M.M.O. is related to the reoccupation of the region in still very cold periods, between MIS 4 and MIS 3; before this, only Mousterian and Taubachian (micro-Mousterian) are present [42]. South of Carpathians, the same broad behaviour is recorded in Kůlna sequence in Moravia [45]. Keilmessergruppen/M.M.O. are also present in more temperate areas, at the margins of their central European core-area, but in particularly cold stages; in south-western France, at Abri du Musée (ensemble V zone II), the Keilmesser with tranchet-blow technique appears together with cold and open environment fauna, like reindeer and mammoth [128]. In eastern France, Grotte de la Vérpiliére II is probably framed between the end of MIS 4 and the beginning of MIS 3, with a quite similar faunal assemblage [40]. The Keilmessergruppen/M.M.O. groups could have been pushed south and west in MIS 4, following large herbivores, when the plains of central Europe were covered by glaciers or permafrost and thus mostly uninhabitable (Fig 24). In these climatic-related population shifts, temporary refuge areas could have been south-western (from Burgundy to Dordogne or Brittany) and south-eastern (Pannonia basin) Europe, before MIS 3 reoccupation [26,106,113]. In this sense, the Central European Keilmessergruppen/M.M.O. settlement systems are thought to be developed in climatically challenged areas of central Europe, between the Alpine and Fenno-Scandinavian glaciers, while classical Mousterian assemblages reflect different strategies of mobility and resources exploitation [129].  Fig 24. Site distribution of Keilmessergruppen techno-complex in the early Weichselian, between MIS 5 and MIS 3. The red zones were possibly populated, the blue zones were occupied by permanent glaciers. In any case, it seems that European Keilmessergruppen/M.M.O. tend to occupy environmentally demanding habitats, often related to stronger environmental constraints including (seasonal) permafrost, long winters and ecological reliance on migratory species. In these contexts, lithic raw materials are difficult to supply, and the Keilmesser could represent an excellent answer due to its long use-life, effectiveness and multi-functionality, good cutting ability, scraping, and butchering of carcasses. Its possible purpose as a core-tool can also imply the production of thin and sharp flakes or scrapers that can serve where the Keilmesser is useless (precision activities or wood-working). This seems to be particularly true for eastern contexts like Crimea, where, however, the climate in MIS 3 was characterized by faint fluctuation between humid/warm and dry/cool intervals, always in boreal forest-steppe landscape [130,131]. In a scenario, the late Middle Paleolithic of Central and Eastern Europe, where “Micoquian option” is generally known, different kinds of sites could be present. Within some open-air, short-term occupations, Keilmesser and related artefacts can represent the whole assemblage, with the absence of core-reduction technologies (i.e. in Lichtenberg [18], Zwolén [31] and Pietraszyn 49a [43]). In these cases, the strategic quality of this artefact as a mobile multi-functional tool is perfectly expressed. Conversely, after longer or more stable occupations, Keilmesser are accompanied by débitage products and were abandoned after more or less long biographies devoted to mobility [9,19,52]. The case of Sesselfelsgrotte G-Complex tells us that Keilmesser-bearing groups are not necessarily characterized by long-distance but frequent mobility patterns, especially in the “initial inventories” where exhausted tools or core-tools are numerous, becoming more logistic and organized during more stable “consecutive inventories” occupations, where resource availability is larger. Furthermore, according to the clear decreasing of strategic multi-purpose tools (core-tools Keilmesser) in relation to the whole backed bifacial tools (only 3 out of 18; Table 10) in the last G1 layer, it is possible that Keilmesser would have gradually increased their value as a cultural symbol and inheritance, rather than their ecological value, that was probably more important for their first conception and diffusion. It is difficult to trace an evolutionary development of Keilmesser shape and function that could give us information in this sense. According to the chronological distinction by Jöris [23], after a debated phase A, the phase B is framed in MIS 4 and characterized by the typical resharpening tranchet blow technique, that could represent a strong cultural marker. Within the MIS 3 phase C this technique would considerably decrease, but this vision is partially revised and corrected [64]. Parallelisms with Sesselfelsgrotte are found in the long sequence of Ciemna cave, where the long-lived Keilmesser are typical of the more ancient layer (layer V), while in a second moment (layer III) these are less numerous and are found together with Levallois products [42]. In both cases, Keilmesser related to “ecological need” (here readable as bifacial core-reduction) would have been typical of a first, developing stage of Keilmessergruppen/M.M.O. In order to advance diachronic interpretations, however, it is necessary to frame this techno-complex and its genesis according to a behavioral point of view. Valoch proposed, as origin of Micoquian assemblages, Eemian contexts like Tata where similar bifacial tools are present [132]. In the same way, the late Acheulean (Jungacheulean) would be at their base for Bosinski [12] and Kozlowski [14], while Jöris [26] asserts that they firstly emerge as a common fund toolkit in different and particular circumstances. The Hypothesis generally depends on whether the long or short chronologies are accepted; however, the early Weichselian (from MIS 5d but especially MIS 4) cooling episodes should be the key moments for the first definition and standardization of these techno-complexes. In these periods, demographic upheavals like displacement and decrease of human groups or mechanisms like bottlenecks may have contributed to form new technical objects or assemblages. Western Europe, in MIS 4, is characterized by the widespread presence of Quina techno-complex, associated to high mobility patterns and exploitation of gregarious and migratory species adapted to harsh environments, like reindeer and bisons [6,133,134]. Lithic technology and economy are aimed at high mobile strategies, resembling Keilmesser-like behaviors; the technical investment is rarely employed to core reduction technologies, but to extend the life length of the flake blanks, here retouched into the typical Quina scrapers. This tool is also a possible matrix from which to detach flakes according to a ramified concept [135–137], thus a mobile blank with strategic value and a rather raw material source. Even if its reduction stages are different if compared to a bifacial tool, at the end of its functional life the Quina scraper may bear bifacial retouching, bulb thinning and triangular shapes [71,135,138,139], being conceptually similar to bifacially shaped tools. The “meeting” between the Quina technology and the Keilmessergruppe/M.M.O. package is well documented, especially in southern Germany, and dated to relatively ancient phases. The main contexts are Bockstein, firstly related to the passage between MIS 5 and MIS 4 [140] but then repositioned between MIS 4 and MIS 3 [69], and of course the most ancient layers of G-Complex in Sesselfelsgrotte, datable at the very beginning of MIS 3 re-occupation of the region after MIS 4. Similarities are noticed with the Rhodanian facies of Quina. Here, in addition to the high rate of retouched tools, tools resembling Keilmesser along with limaces are present and in contexts datable to the beginning of MIS 3 and at the limits of Keilmessergruppen/M.M.O. influence area [123]. Therefore, in this time span and in an area yet to be defined, a meeting and a partial coexistence of these two concepts have occurred. In the first occupational cycles of Sesselfelsgrotte G-Complex, the production of unifacial and bifacial blanks follow the same notions, that are the application of Quina concepts [9]. In the following phases, instead, there is the clear distinction between the core-reduction methods (mainly Levallois) for the production of flakes, and the manufacturing of simple or plano-covex bifacial tools on the other side. In Bockstein, the same behavior is documented, wherein small “bockstein-messer” are manufactured on Quina blanks [69]. This could represent the techno-functional integration of a tool-type, the backed knife, that in classical Quina assemblages is very differently shaped [70]. In a preliminary way, we can hypothesize that the first conception and production of the Keilmesser tool could have derived from ecological adaptations, which resulted in a series of actions functional to practical objectives. In this case, the aim was probably the manufacturing of a highly potential, multifunctional backed knife. Later in time, these tools could have become a cultural symbol, in a culturally and technologically fragmented Late Middle Paleolithic Europe, characterized by widespread regionalisms. Probably due to climatic and demographical shifts faced by late Neanderthal groups in the first Weichselian glacial maximum, a very diverse cultural mosaic is shaped and archaeologically perceptible [10]. In these phases the tool is more likely the result of the achievement of a conceptual scheme and, perhaps, equipped with aesthetic and symbolic value. The cultural and possibly ethnic value of these industries has recently been reinforced with data from the early-MIS 3 Neanderthals from Chagyrskaya in the Altai mountains: here, techno-typological affinities with Central- and Eastern-European Micoquian come together with genetic similarities with European specimens, while the same groups were unquestionably different from earlier Altai Neanderthals bearing Mousterian assemblages [141,142]. 8. Conclusions The techno-functional and morphological revision of backed implements from Sesselfelsgrotte G-complex allowed a broader understanding of the relationship between the different techno-typological entities. The main conclusions achieved through this work are the following: The Keilmesser tool-type is not always well defined but actually quite fluid. On a techno-functional base it includes several sub-types implying distinct functional operational schemes according to morphology, functional units, dimensions, etc. These schemes could both have direct functional or reductional values, since some of them (especially the pointed implements) could have represented the initial schemes, that eventually changed due to resharpening and retooling. This variability is confirmed by past use-wear analyses, perfectly showing the versatility of this concept. An approach that proved to be particularly useful is the working-step analysis applied to techno-functional schemes, able to discern 3 types of Keilmesser distinct in their ecological biographies and potentialities. Among these, the core-tools recorded their multi-purpose concept as raw material matrix and asymmetrical backed tool. Enlarging the analyses to the other backed implements, including backed scrapers or simple flake-products, it is evident that a certain degree of overlapping affects these typological categories, firstly evident within 3D shape analyses. Even if some preferences are recorded among techno-functional schemes, no exclusive scheme for a single tool type exists. Moreover, comparisons between cutting edges’ angles and active bevel analyzed in virtual section proved that Keilmesser’s plano-convex section was probably manufactured in order to imitate the functionality of flake tools, besides the applicability of angle mainteinance techniques. Lastly, the backing type is related more to the initial blank than to the tool type. Past use-wear analyses provided consistent data in this sense, since typology is never associated automathically to a function, and backed knives are found among very different kinds of tools. The main discriminating factor between backed bifacial tools and unifacial tools is therefore the higher potential and use-life length of the first tools. For these reasons Keilmesser are the ideal strategic backed implements, mainly hand-held and useful for cutting, scraping and occasionally to produce smaller flakes from their flat surface. These tools and the so-called “Micoquian option” they represent are deeply interrelated to the Mousterian techno-tipological substrate and emerge in constrained ecological conditions related to patterns of frequent mobility. These conditions favored the production of highly strategic, long-life blanks and the ramification of knapping reduction sequences. In the absence of large-scale reliable seasonal data on the occupations, we can assert that possibly major climatic shifts are at the base of their emergence, framed around the end of MIS 5 and the beginning of MIS 3. Broad demographic dynamics, following the expansion and contraction of ice covering and related herds of herbivores, resulted in a widespread techno-cultural regionalization from which also keilmessergruppen/M.M.O. emerged. This is why Keilmesser, after their initial ecological and strategic signficance, could have then represented a tool equipped with cultural-related meaning. Sesselfelsgrotte G-complex seem to weakly document this kind of behavior. However, a broad comparison between early and late Keilmessergruppen/M.M.O. contexts is needed in order to verify this claim. Moreover, extensive chronometrical analyses will clarify the reliability of the long or short chronologies applied to Keilmessergruppen/M.M.O. Future perspectives, in this sense, may also come from a more intense and targeted experimental protocol aimed at measuring the efficiency of Keilmesser in relation to other similar backed tools typical of other techno-complexes, in order to further quantify the ecological efficiency or the presumed cutural valence. The backing concept, here embedded in the raw blank use strategies, is particularly important in this chrono-cultural phase, when intentional backing by means of retouch appears in relation to specialized and precision tools [75]. In this sense, the bifacial shaping of back’s angles and conjunctions seems to be a peculiar technical behavior, not documented elsewhere, that deserves an indepth investigation. The current research approach evaluates the technical objects in relation to their usefulness, environmental adaptation and also cognitive factors such as lateralization and planning depth. When this relationship isn’t clear or is lacking, traditions and fashion may have played a more important role, especially in Late Middle Paleolithic Europe, fragmented into a technological, cultural and possibly biological mosaic. |
|
|
|
Post by Admin on Sept 30, 2020 20:21:12 GMT
The early Aurignacian dispersal of modern humans into westernmost Eurasia PNAS first published September 28, 2020 doi.org/10.1073/pnas.2016062117Edited by Richard G. Klein, Stanford University, Stanford, CA, and approved August 26, 2020 (received for review July 30, 2020) Significance We report the remarkable discovery of an early Aurignacian occupation, ∼5,000 years older than any Upper Paleolithic site in westernmost Eurasia. The archaeological and radiocarbon data provide definitive evidence that modern humans were in western Iberia at a time when, if present at all, Neanderthal populations would have been extremely sparse. This discovery has important ramifications for our understanding of the process of modern human dispersal and replacement of Neanderthal populations. The results support a very rapid, unimpeded dispersal of modern humans across western Eurasia and support the notion that climate and environmental change played a significant role in this process. Abstract Documenting the first appearance of modern humans in a given region is key to understanding the dispersal process and the replacement or assimilation of indigenous human populations such as the Neanderthals. The Iberian Peninsula was the last refuge of Neanderthal populations as modern humans advanced across Eurasia. Here we present evidence of an early Aurignacian occupation at Lapa do Picareiro in central Portugal. Diagnostic artifacts were found in a sealed stratigraphic layer dated 41.1 to 38.1 ka cal BP, documenting a modern human presence on the western margin of Iberia ∼5,000 years earlier than previously known. The data indicate a rapid modern human dispersal across southern Europe, reaching the westernmost edge where Neanderthals were thought to persist. The results support the notion of a mosaic process of modern human dispersal and replacement of indigenous Neanderthal populations. Aurignacianmodern humandispersalIberiaPaleolithic The dispersal of modern humans across western Eurasia, associated with the Upper Paleolithic, is well documented compared to other regions but still susceptible to discoveries that can overturn prevailing ideas, especially those based on first appearance dates (1, 2). Current data support an east-west dispersal beginning ∼46 ka cal BP in the Balkan Peninsula at Bacho Kiro (3, 4) (Fig. 1). Subsequently, modern humans spread up the Danube river basin and along the Mediterranean rim within a relatively short period (5). The process was likely a mosaic involving dispersal into empty spaces and interaction with indigenous Neanderthal populations. At some point, around 43 to 42 ka cal BP, the regional variants of the Initial Upper Paleolithic coalesced into the Aurignacian technocomplex, appearing synchronously across western Eurasia (6).  Fig. 1. Map of selected Early Upper Paleolithic (red and pink) and Late Middle Paleolithic (blue) sites in Europe (7). (1) Arbreda, (2) Reclau Viver, (3) Abric Romaní, (4) Cova Gran, (5) Mallaetes, (6) Cova Foradada, (7) Cova de les Cendres, (8) Sima de las Palomas, (9) Cova Beneito, (10) La Boja, (11) Cueva Antón, (12) Zafarraya, (13) Bajondillo, (14) Gorham’s Cave, (15) Pego do Diabo, (16) Mira Nascente, (17) Lapa do Picareiro, (18) Gruta da Oliveira, (19) Foz do Enxarrique, (20) Cardina, (21) A Valiña, (22) La Viña, (23) El Sidrón, (24) El Castillo, (25) Cueva Morín, (26) El Mirón, (27) Labeko Koba, (28) Aitzbitarte III, (29) Isturitz, (30) Abri Castanet, (31), Abri Pataud, (32) Les Cottés, (33) Kent’s Cavern, (34) Riparo Mochi, (35) Geissenklosterle, (36) Fumane, (37) Willendorf, (38) Grotta del Cavallo, (39) Pestera cu Oase, and (40) Bacho Kiro. The Iberian Peninsula holds a peculiar place in the problem of modern human dispersal ever since the publication of unexpectedly early dates for the first Upper Paleolithic appearance at El Castillo, l’Arbreda, and Abric Romaní in northern Spain (8, 9). Subsequent dating from these and additional sites has constrained the Aurignacian arrival in the region to ∼43.3 to 40.5 ka cal BP (10⇓⇓⇓–14). These dates verify a rapid modern human dispersal and allow for a temporal overlap of ∼1,000 years with Neanderthals in northern Spain (11), and longer in southern Iberia. Furthermore, the scenario is complicated by the lack of associated fossil remains, leaving open the possibility that either human group created the early Aurignacian assemblages in the Franco-Cantabrian region (15). Despite this lack of direct association for the early Aurignacian, modern human remains have been identified in this time frame in Romania (16) and Italy (17, 18). Since no Neanderthal sites contain evidence for the use of carinated technology to produce bladelets, we can assume that modern humans were the makers of the entire Aurignacian cultural complex. Undeterred by this uncertainty, the early appearance dates for the Upper Paleolithic and late appearance dates for Middle Paleolithic Neanderthals in southern Iberia led to the construction of various models to explain the apparent biogeographic boundary separating the two populations (19, 20). In these models, ecological adaptations allowed Neanderthals to survive, preventing modern human dispersal in southern Iberia until ∼37 to 30 ka cal BP, a period of 6,000 to 12,000 years (21, 22). The recent dating of Bajondillo cave, on the southern coast of Spain, arguably demonstrated the first presence of modern humans at ∼45 to 43 ka cal BP, suggesting an even more expansive dispersal across Europe in a geological blink of an eye (23). The new Bajondillo dates pushed the first appearance of modern humans several thousand years earlier in time, upsetting previously held views. Critics dismissed the dated lithic assemblage in level Bj13 as a mixed collection of artifacts from younger and older occupations or as lacking typological traits of the Proto or Early Aurignacian technocomplexes (24, 25). Until the Bajondillo publication, the earliest Upper Paleolithic held at ∼35 ka cal BP at Cova de les Cendres (Mediterranean coast) (26), ∼36.5 ka cal BP, at La Boja (22), (southern Spain), and ∼34.5 ka cal BP at Pego do Diabo, (central Portugal) (27), all attributed to the Evolved/Late Aurignacian. On the flip side of the problem, the last appearance of Neanderthals and the Middle Paleolithic has also been cast in doubt (28, 29). The number of “late” Neanderthal sites (<42 ka cal BP) has diminished substantially since the application of new dating techniques showed them to be much older (>42 ka cal BP) than previously thought (30⇓⇓–33). Across central Spain, there are no Neanderthal remains or Middle Paleolithic sites dated after 42 ka cal BP (34⇓–36). At present, Gruta da Oliveira, Cueva Antón, and Gorham’s Cave remain the only “late” Neanderthal sites in southern Iberia dated ∼37 ka cal BP or later (21, 22). Clearly, sites dated ∼42 to 37 ka cal BP are extremely rare, suggesting that most of Iberia south of the Ebro basin was a sparsely, if at all, populated landscape into which modern humans could have easily dispersed. The lack of archaeological and fossil evidence for this interval could also be due to climatic and landscape instability that erased the record or prevented its formation (37, 38). Under these conditions, survivorship is likeliest in sedimentary traps or sheltered locations where accumulative processes preserve material evidence. One such place is Lapa do Picareiro, a limestone cave located 570 m above sea level, on the west-facing slope of Serra de Aire, a karst mountain north of the Tagus River valley in west-central Portugal (Fig. 1). The 15 × 15 m cavern (Fig. 2 A and B and SI Appendix, Fig. S1) is part of a large (25 × 30 m) bedrock depression with a thick sedimentary fill of muddy éboulis representing much of the Late Pleistocene. We have excavated a 10.6-m deep section with 36 Pleistocene-aged strata (levels E-NN), revealing a thick Upper Paleolithic sequence (levels E-II) continuing into the Middle Paleolithic (levels JJ-NN) (39, 40). Age determination of the levels comes from 80 radiocarbon dates produced over the last 25 y of investigation at the cave. The stratigraphic sequence in Picareiro has roughly 2 m of sediment dated ∼45 to 35 ka cal BP, corresponding to the temporal range of the Middle-Upper Paleolithic transition in southern Iberia (Fig. 2 C–E and SI Appendix, Figs. S2 and S3). As the ongoing excavation has exposed the deeper deposits in the back of the cave, evidence for previously undetected Early Upper Paleolithic occupations has emerged.  Fig. 2. (A) Site plan with profile locations shown in red. (B) Orthophoto of the excavation from above. (C) Stratigraphic profiles for the MP-UP sequence in Lapa do Picareiro. (D) Level X to level II profile in units B8-A8, (B) level GG to level JJ in unit F9, (E) close-up of level GG-II in unit C8. |
|
|
|
Post by Admin on Sept 30, 2020 23:24:39 GMT
Results A series of dates using the ultrafiltration and enhanced collagen extraction pretreatments on anthropically modified ungulate bones from the Late Middle and Early Upper Paleolithic levels is presented here (SI Appendix, Table S1 and Figs. S4–S6). The results confirm an early Aurignacian presence in the region, potentially overlapping with level Bj13 from Bajondillo and positing significant implications for our understanding of modern human dispersal and late survival of Neanderthals in Europe. Fig. 3 shows the plots of stratigraphically distinct lithic artifacts with associated radiocarbon dates. The levels display a high degree of lithic assemblage integrity supported by the technological characteristics, raw material representation, and systematic artifact refitting. The radiocarbon dates also reflect stratigraphic integrity with no significant inversions between the levels. Furthermore, the dated bone samples have fresh, well-preserved green bone fractures with no signs of trampling or abrasion (SI Appendix, Figs. S4–S6) and were taken from deposits with a low (2°–4°) inclination (40). The sedimentological and geochemical details published in Benedetti et al. (40) provide additional support for the integrity of the deposits, which are evidenced by distinct beds, varying from roughly 5 to 30 cm thick, with uniform properties of clast size and fine sediment content. Thus, vertical and nonvertical migration in the matrix appears to have been negligible. The dates reported here are included with previous determinations in a Bayesian model constructed using the new IntCal20 in the OxCal program 4.4 (41, 42) (Fig. 4 and SI Appendix, Table S2). The temporal gaps between the layers reflect the sampling of archaeological horizons within them, and the slow rate of sedimentation reported previously (40).  Fig. 3. Plots of lithic artifacts along with radiocarbon-dated bone samples from Lapa do Picareiro: (A) X-axis profile of the cave, perpendicular to the central axis, (B) Y-axis profile of the cave, lengthwise from front to back.  Fig. 4. Bayesian model for the Lapa do Picareiro. Radiocarbon dates are calibrated using IntCal20 (41); the model and boundaries were calculated using OxCal 4.3 (42), including a general t-type outlier model. Outlier prior and posterior probability are shown in square brackets. Four samples are excluded from the model iterations by giving them a prior outlier probability of 100% because of the poor collagen preservation. The chronology is compared to the North Greenland Ice Core Project (NGRIP) Greenland Ice Core Chronology 2005 (GICC05) (43) δ18O paleo-environmental record with Greenland Interstadials (GI) 12 to 3 and Heinrich events (H) 5, 4, and 3 indicated. The base of the sequence discussed here, level JJ, is a ∼1-m-thick layer of small-medium limestone clasts with reddish-brown fine sediment (SI Appendix, Table S3). The deposit contains lithic artifacts made using discoidal core/flake technology typical of the Middle Paleolithic, dispersed charcoal, and animal bones. The radiocarbon samples were taken from two artifact horizons separated by a ∼20-cm-thick dark sediment lens. The lower horizon dates to ∼51.5 to 44.1 ka cal BP and the upper horizon dates to ∼45.0 to 43.5 ka cal BP. The top 20 to 30 cm of level JJ, dated 42.9 to 42.4 ka cal BP, contains bones with percussion marks consistent with intentional butchery by humans but lithic artifacts have not yet been found in this zone (Fig. 5D).  Fig. 5. Lithic artifacts from the Middle to Upper Paleolithic transition levels at Lapa do Picareiro. (A) Chert cores and core tablet from level DD. (B) Quartzite flakes from level FF. (C) Early Aurignacian carinated endscrapers/cores and bladelets from level GG. (D) Middle Paleolithic core and flakes. Levels II through GG represent a ∼40-cm-thick, distinct, and partially cemented portion of the sequence that caps level JJ. The lower part, level II is a layer of medium and larger clasts with brown mud cemented by calcite followed by a layer of medium-sized clasts with brown mud that is uncemented, level HH. Level GG is a mostly cemented layer of larger limestone clasts and brown fine sediment that extends across the center of the cave. In the back of the cave, the calcite cement gradually disappears. The thick, firmly cemented parts of level GG in the center of the cave appear to have formed by postdepositional precipitation from calcium-enriched water penetrating the interstitial spaces between limestone clasts on the cave floor (SI Appendix, Fig. S4). A sharp peak in magnetic susceptibility values recorded in level II indicates a brief warm period followed by low values in levels HH and GG, corresponding to a period of prolonged cold, dry conditions during its formation (40). The precipitation of carbonate cement likely occurred at the end of this cold episode with the return of humid conditions. The earliest Aurignacian artifacts are distributed throughout the muddy matrix from the base of the large clasts of level GG through level II (Fig. 2 C–E and SI Appendix, Fig. S7). The radiocarbon samples were taken from the uncemented areas of level GG and II, in direct association with the levels GG-II artifacts. Levels GG-II contain a diagnostically Upper Paleolithic lithic assemblage comprised of small bladelets and carinated endscrapers (now recognized as cores) typical of the early Aurignacian (6, 44, 45) (Fig. 5C). The bladelets have the characteristic dimensions and shape, lacking retouch, similar to those found in the early Aurignacian assemblages of northern Iberia (46). This assemblage is made primarily from chert but also includes a small number of quartz flakes. Accelerator mass spectrometry (AMS) radiocarbon dating of anthropically modified, medium-sized ungulate bones provide bracketing ages of ∼41.9 to 41.1 ka cal BP (terminus post quem or earliest possible date) and 39.4 to 38.1 ka cal BP (terminus ante quem or latest possible date). Thus, the assemblage falls within most of the Proto-Aurignacian and entirely within the Early-Aurignacian time frame in Europe (11, 47, 48). The artifact deposition likely took place during the Greenland Stadial (GS)-9 climate phase associated with Heinrich Event 4 (H4) (39.9 to 38.2 ka) or GS-10 (40.8 to 40.1 ka), but possibly even earlier during the time of Greenland Interstadial (GI)-10 (41.4 to 40.8 ka) and GS-11 (42.2 to 41.5 ka) (43). Most of the artifacts plot within a ∼20-cm linear band between the dated samples, making a more exact age estimation or paleoclimatic association difficult at this time, but these still represent the oldest, most precise, and reliable dates for the Aurignacian in western Iberia. In stark contrast, overlying sediments of level FF are composed of loose, small-medium limestone clasts and dark reddish-brown fine sediment. This layer contains undiagnostic quartz and quartzite flakes, animal bones, and dispersed charcoal dated 38.6 to 36.4 ka cal BP (Fig. 5B). The organic matter content and high magnetic susceptibility values in level FF reflect a period of relatively mild climate associated with GI-8 (43). Level EE is a thin layer (5 to 10 cm) of small clasts with reddish-brown fine sediment that appears to be archaeologically sterile. The radiocarbon date for level EE (36.7 to 36.1 ka cal BP) is indistinguishable from those of level FF. The dated bone sample plots in line with those from level FF and, therefore, the MAMS-44444 (Mannheim AMS lab at the Curt-Engelhorn-Centre for Archaeometry) specimen may not accurately date level EE. Level DD is a 20- to 30-cm-thick layer of medium-large limestone clasts with brown fine sediment. This layer, dated 35.4 to 34.8 ka cal BP, contains a lithic assemblage almost exclusively on chert, characterized by the production of large flakes using prismatic core technology (Fig. 5A). The dates for these levels place them chronologically within the Aurignacian time frame in western Europe (47, 48) but the assemblages lack diagnostic pieces for a particular phase. Low magnetic susceptibility values and radiocarbon dates correspond to GS-8 (43). Despite their small size, comparison of the lithic assemblages reveals important differences in the frequencies of the different classes of blanks across all levels (SI Appendix, Table S4). While levels JJ, FF, and DD are dominated by complete flakes and flake fragments, the level GG-II assemblage is marked by a high frequency of bladelets and bladelet fragments. Blades are also present in the level GG-II assemblage but completely absent from all of the remaining levels assessed here. Overall, cores and retouched tools are very rare in all assemblages (SI Appendix, Table S5). The levels also differ in lithic reduction strategies. In level JJ, centripetal, subcentripetal, and other reduction patterns are equally represented, typical of a Mousterian assemblage, while for all of the overlaying levels unidirectional strategies are predominant. Raw material use is also markedly different across levels (SI Appendix, Fig. S8). Levels JJ and FF are characterized by the use of quartzite and quartz, with chert representing only 15% in the Mousterian level JJ, and completely absent from level FF. In levels GG-II and DD, the scenario is totally different as chert is the most frequent raw material (>75% of all artifacts), with milky quartz/rock crystal (in GG-II) and quartzite (in DD) making up the rest of the assemblages. A noteworthy number (n = 12) of chert artifacts in level DD refit into four different sets, revealing both the integrity of the assemblage and the occurrence of onsite knapping activities. While no refits have been made among the chert bladelets and the carinated endscrapers recovered in level GG-II, comparison between the bladelets and the last scar on the flaking surface of the carinated elements suggest a single reduction sequence for producing small bladelets (SI Appendix, Table S6). Abundant faunal remains have been recovered in all of the levels presented here. Preliminary results of ongoing analyses indicate that large and small mammal taxonomic representation changed very little across the Late Middle and Early Upper Paleolithic layers. SI Appendix, Tables S7–S10 provide results of taxonomic identifications made to date. Large and small mammals for all levels include red deer, ibex, and rabbit. Some of the ungulate remains show evidence for butchery with cut marks, percussion scars, and long bone fractures consistent with marrow removal. Horse was also exploited in the Late Middle Paleolithic level JJ. The rabbits show little if any direct evidence for human exploitation but only a very small proportion of the assemblage has been analyzed in detail. Carnivores are primarily represented by lynx in levels DD-FF and JJ. Fox also appears in the Late Middle Paleolithic. A variety of bird taxa are also present in the assemblages, but it is not clear if the remains were brought to the cave by humans. Ongoing taphonomic analyses should resolve this issue. The same can be said for the micromammal and herpetological taxa. |
|
|
|
Post by Admin on Oct 1, 2020 4:12:36 GMT
Discussion
The stratigraphy, techno-typological analysis of the lithic artifacts, and radiocarbon dating demonstrate that the level GG-II assemblage represents a discrete occupation layer, wholly distinct from those in the levels above and below. The assemblage is small but consistent with the attributes of the early Aurignacian. The dates presented here conservatively place the level GG-II occupation at ∼41.1 to 38.1 ka cal BP.
Our results from Picareiro provide definitive evidence that modern humans were in western Iberia at a time when, if present at all, Neanderthal populations would have been extremely sparse. Our data offer some resolution to the implications of Wood et al. (29) scenarios for the ∼42 to 37 ka cal BP time frame. We can discount their scenario in which southern Iberia was abandoned by both Neanderthals and modern humans and confirm the one in which modern humans spread into the southern regions soon after they arrived in northern Iberia. As for the scenario in which Neanderthals were present in southern Iberia until very late, the Picareiro data alone cannot resolve this issue. Middle Paleolithic occupations at Picareiro ended by 42.5 ka cal BP, but they apparently continued until ∼36 ka cal BP at Gruta da Oliveira just 4.2 km away, based on current evidence (49, 50).
Our results also cast further doubt on the idea of a “hard border” or frontier between Neanderthal and modern human populations between ∼42 and 37 ka cal BP (22, 51). Instead, the Ebro river valley was likely a permeable landscape feature that facilitated dispersal (52). Modern humans may have encountered a few remnant Neanderthal groups but it appears that most of Iberia south of the Ebro was already depopulated (35). This pattern is evident in Portugal, where Middle Paleolithic end dates cluster at ∼45 to 42 ka at Foz do Enxarrique (30), Mira Nascente (53), Lapa do Picareiro, and Cardina (54), followed by a nearly total absence of evidence for Neanderthals on the landscape.
The Picareiro data confirm a rapid modern human dispersal across Iberia that opens up additional lines of inquiry, testable hypotheses, and explanatory scenarios for the Middle to Upper Paleolithic transition. First, the early Aurignacian arrival indicates a substantial time gap between the Picareiro record of Early Upper Paleolithic occupations and the rest of the region (27, 54). This may reflect an expansion of small pioneer groups that did not leave a highly visible footprint due to low population density (55) or did not establish a permanent foothold in the region. Alternatively, climate-induced erosive episodes may have erased much of the archaeological evidence of their presence on the landscape (37, 38). Either case limits the detectability of the earliest pioneers. Another possibility is that the evidence exists among assemblages in the region lacking diagnostic elements and/or radiometric dates. Picareiro level FF, dated within the Evolved Aurignacian time frame, exemplifies this case with simple flakes, mainly quartzite and quartz, and no formal tools. Aside from its stratigraphic position and radiocarbon dating, there is nothing diagnostically Aurignacian about the assemblage. Level FF represents a low-cost, expedient technology often attributed to the Middle Paleolithic in Iberian sites dated ∼42 to 32 ka cal BP (56). Assemblages like these may have been a regular element of the pioneer phase in modern human dispersal (55). Just how widespread and common they were during the Aurignacian time frame remains to be investigated but similar expedient core reduction strategies are known throughout the Upper Paleolithic at many sites (57), including Picareiro.
Second, the successive climatic downturns between 44 and 40 ka may have created new opportunities for modern human dispersal into the region, as predicted by Banks et al. (47), possibly following the southward range expansion of familiar Eurosiberian taxa along the Atlantic margin or the major east-west river drainages such as the Duero or Tagus (SI Appendix, Fig. S9). River systems like these played a key role as “communication corridors and mobility conduits” for the dispersal of modern humans across Europe (52). A critical aspect of dispersal would have been the development of cognitive maps to navigate unknown landscapes, and rivers are the easiest spatial features to follow (58). New optically-stimulated luminescence (OSL) ages for Cardina, an open-air site in the Douro drainage basin of northeast Portugal, show a long hiatus between the last Middle Paleolithic occupation ∼42.9 ka and the Evolved Aurignacian occupation ∼33.6 ka (54). The absence of an earlier Aurignacian occupation could rule out the Douro valley as a dispersal route, but the area may eventually yield supportive evidence. Rather, the spread of modern humans across the Iberian Peninsula may represent a “jump dispersal” through which they avoided or bypassed unproductive or high-risk areas like the more arid interior of Iberia (2). Along interior river drainages, rapid dispersal may have been necessary to mitigate water scarcity during extreme droughts associated with Heinrich events, thus explaining the lack of early Aurignacian sites. Alternatively, the coastal route hypothesis advanced by Cortés-Sánchez et al. (23) is also supported by the distribution of Early Upper Paleolithic sites along the Iberian coast (59). This ecotonal position likely provided more predictable resources and less ecological risk during periods of climatic stress. The topography of coasts would have also facilitated communication and transmission of information among pioneer groups (55).
Third, Neanderthals and modern humans may have been contemporary and in close proximity in the limestone massif of Estremadura in Portugal. If so, there is no evidence that they were in direct contact as the Picareiro GG-II occupation took place between the occupations of Gruta da Oliveira levels 9 and 8. However, level FF, without diagnostic artifacts, is contemporary with Oliveira level 8 and could indicate either coexistence or successive, alternating presence of different populations. The presence of modern humans overlapping in time lends support to competitive exclusion as an explanation for Neanderthal extinction (60, 61). On the other hand, if the Gruta da Oliveira level 8 dates are erroneously too young, as postulated by Wood et al. (29), then there are no Neanderthal or Middle Paleolithic sites in Portugal that postdate ∼42 ka cal BP. Thus, there may be no temporal overlap or competition between the last Neanderthals and earliest modern humans in the region.
Lastly, the Picareiro record appears to reflect the pattern across much of western Eurasia where sterile layers between the last Neanderthal and modern human occupations have been linked to millennial-scale climate cycles and environmental change (62). Depopulation appears to have occurred during severe cold and dry stadials that disrupted and fragmented habitat patches across western Eurasia, negatively impacting Neanderthal populations and opening new spaces for modern human dispersal (63). In Iberia, paleoclimate records show regional variability in the terrestrial response to GS-12 (44.3 to 43.3 ka), GS-11 (42.2 to 41.5 ka), and GS-9 or HS-4 (39.9 to 38.2 ka) (64, 65). The archaeological and sedimentological data from Picareiro and other sites are too coarse grained at present, but the timing of these successive perturbations appear to coincide with regional Neanderthal depopulation. At Picareiro, the last Middle Paleolithic occupation corresponds to the beginning of GS-12, followed by an apparent occupational hiatus in the upper 20 to 30 cm of level JJ, with the subsequent Aurignacian arrival between GS-11 and GS-9.
Although we may never know the first actual presence (2) of modern humans or the last actual presence of Neanderthals in Iberia, south of the Ebro basin, the data from Lapa do Picareiro expand our knowledge about the dispersal of modern humans. Based on current understanding of Middle and Upper Paleolithic technological associations, the Picareiro case provides definitive evidence for the early appearance of modern humans in westernmost Eurasia, disrupting previous models and creating opportunities for new lines of inquiry. A major gap in our knowledge of the 42 to 37 ka cal BP interval remains to be filled with further investigation and continued field work.
|
|
|
|
Post by Admin on Oct 8, 2020 22:27:33 GMT
Early development of the Neanderthal ribcage reveals a different body shape at birth compared to modern humans Science Advances 07 Oct 2020: Vol. 6, no. 41, eabb4377 DOI: 10.1126/sciadv.abb4377 Abstract Ontogenetic studies provide clues for understanding important paleobiological aspects of extinct species. When compared to that of modern humans, the adult Neanderthal thorax was shorter, deeper, and wider. This is related to the wide Neanderthal body and is consistent with their hypothetical large requirements for energy and oxygen. Whether these differences were already established at birth or appeared later during development is unknown. To delve into this question, we use virtual reconstruction tools and geometric morphometrics to recover the 3D morphology of the ribcages of four Neanderthal individuals from birth to around 3 years old: Mezmaiskaya 1, Le Moustier 2, Dederiyeh 1, and Roc de Marsal. Our results indicate that the comparatively deep and short ribcage of the Neanderthals was already present at birth, as were other skeletal species-specific traits. This morphology possibly represents the plesiomorphic condition shared with Homo erectus, and it is likely linked to large energetic requirements. INTRODUCTION Prenatal and early postnatal growth and development are crucial to understanding the adult size and shape of the different anatomical regions because of the large number and high rate of size and shape changes occurring in the human body during those phases (1–5). Also, from an evolutionary point of view, prenatal and early postnatal ontogeny are decisive because evolution happens via phylogenetic modification of the ontogenetic processes that occur mostly in those phases (3, 6, 7). 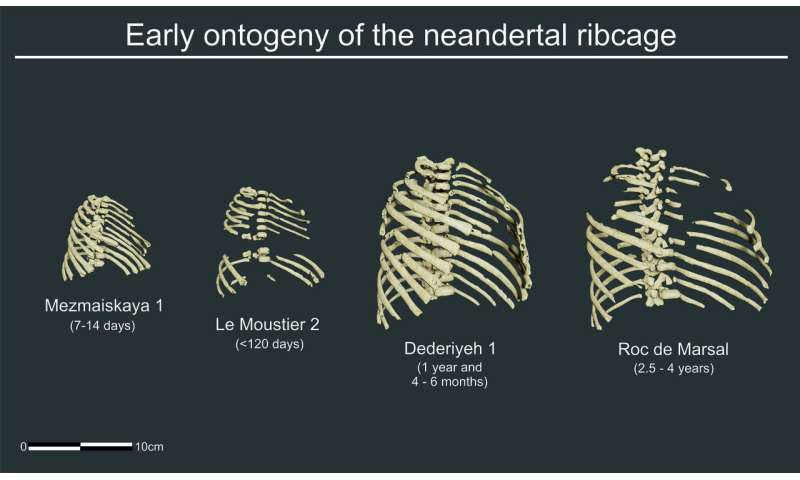 Adult morphologies can vary because of interspecific differences in the shape of an anatomical element at the moment of birth that are caused by differences in the prenatal ontogenetic trajectories or because of differences in the shape of an anatomical element that arise after birth that are caused by differences in the postnatal ontogenetic trajectories, either concerning their orientations, lengths, or a combination of both (1). Roughly speaking, if morphological differences are found at birth and the postnatal ontogenetic pattern is equal in the two species, their ontogenetic trajectories will be parallel. Conversely, if they have a similar morphology at birth but show differences in the postnatal ontogenetic pattern, their ontogenetic trajectories will be divergent (1–3). This distinction is important because parallel postnatal ontogenetic trajectories between two closely related species could point to a consistency of genetic regulation of that anatomical element (1). In addition, the fact that a morphological feature is already present at birth will suggest that it is a relevant taxonomical characteristic not caused by developmental plasticity. Ontogenetic trajectories in Neanderthals Despite genetic similarities that allowed for admixture (8), there is a well-established consensus that Neanderthals showed significant morphological differences when compared to modern humans (MHs) in the cranium and postcranium (9, 10). Some of these differences are plesiomorphic inherited traits from their Early or Middle Pleistocene ancestors, while others are present exclusively in Neanderthals (autapomorphies) (11, 12). Neanderthals were highly encephalized (4, 13, 14) and heavy-bodied hominins (15, 16) requiring large amounts of energy (17–19). It has been proposed that to fulfill these energetic demands, the Neanderthal thorax had a large estimated total lung capacity (19) and a different thoracic shape that included a shorter, slightly deeper, and mediolaterally larger chest with more horizontally oriented ribs and a more invaginated thoracic spine, compared to MH (19–26). The very specific Neanderthal traits found throughout the skeleton (i.e., those different in size and shape from MH) are the result of differences present at birth and/or differences in the postnatal ontogenetic pattern, which may vary in different skeletal regions. However, despite being the best-known extinct human species, there are only a few studies on the Neanderthal postnatal ontogeny due to the paucity of well-preserved subadult fossil remains, especially of the postcranium. Nonetheless, despite the limited record, some patterns have been proposed, providing evolutionary insights. For example, MH and Neanderthal femoral length followed similar growth patterns with no differences at birth (27). Other anatomical traits (e.g., general cranium shape, clavicle length, and femoral and tibial robusticity) seemed to be different at birth between the two species and followed parallel ontogenetic trajectories, resulting in different adult shapes (2, 27, 28). Last, in the case of the mandible (2, 29) and the brain (4, 13, 14), Neanderthals and MH had not only different shapes at birth but also divergent growth patterns. However, there are still many anatomical regions that are relatively well known in the Neanderthal adult record for which there are few ontogenetic studies, which is the case of the thorax (24, 25). Methodological improvements in virtual reconstruction and statistical missing data estimation have improved the knowledge of the adult Neanderthal thorax (26). However, ribs and vertebrae from perinates and infants are smaller and more fragile, which represents a major challenge during the study of the early postnatal ontogeny of the Neanderthal thorax. So far, only basic descriptions and inventories of fossil ribs and vertebrae have been available (30, 31), and artistic license was used when ribcage reconstructions of subadults were made (4). Apart from this very basic knowledge, the little information we have about this issue comes from (i) descriptive anatomy of the prenatal (32) and early postnatal ontogeny of MH (33, 34) and (ii) late postnatal ontogeny of the Neanderthal first ribs (20). Research on prenatal ontogeny of the MH ribcage has found that all thoracic dimensions (anteroposterior, craniocaudal, and mediolateral) are modified during the fetal period to result in the newborn ribcage (32). All these dimensions develop differently in the different rib levels: For example, all levels have roughly the same anteroposterior relative length in early fetuses, whereas the upper and central ribs of late fetuses are much deeper, relatively, than the lower levels (32). This is consistent with research on later postnatal ontogeny of the human ribcage, which has found that, after birth, the upper and lower thorax have a differential development that gives rise to the adult ribcage of MH, which is relatively expanded in the cranial part and narrow in the caudal part (33, 34). This differential development, controlled by Hox gene expression (35), is crucial because it indicates that slight modifications during development at different rib levels would cause different ribcage morphologies. This could have evolutionary implications for understanding the adult thorax not only in our own species but also in other hominins such as Neanderthals. In addition, the only study that tackled the postnatal ontogeny of the thoracic skeleton in this species was carried out by Bastir et al. (20). They found divergent ontogenetic trajectories in the first ribs of MH and Neanderthals, the latter showing less curved first ribs in the youngest specimen (La Ferrassie 6) and along the entire postnatal ontogeny when compared to MH. However, we do not know to what extent this could be extrapolated to the entire thorax. In this study, we used virtual and statistical methods to reconstruct the ribcage of four young Neanderthal specimens (Table 1), identifying potential differences with MH in thorax morphology affecting the evolution of body shape and influencing respiration. Specifically, we reconstructed the ribcages of perinatal individuals of Mezmaiskaya 1 [M1; 7 to 14 days (4)] and Le Moustier 2 [LM2; <120 days (36)] and infant individuals from Dederiyeh 1 [D1; 1.41 years (37)] and Roc de Marsal (RdM; 2.54 years (31)]. We also provided the first three-dimensional (3D) morphological assessment of the early postnatal ontogeny of the MH ribcage during the decisive first 3 years of postnatal life to serve as a comparative baseline. Because of the differences in this anatomical region in adults, we tested whether Neanderthal thorax morphology was already different from that of MH at birth. |
|

