|
|
Post by Admin on Aug 30, 2022 21:15:18 GMT
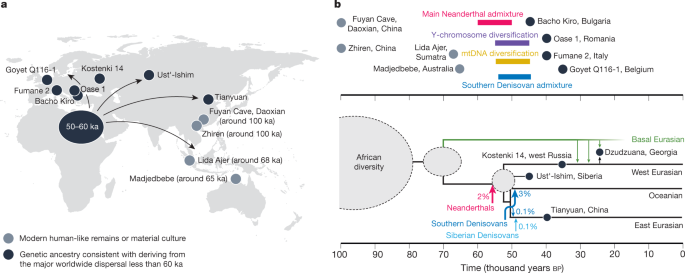 Neanderthal admixture35,46, around 45–55ka. Like Neanderthal ances-try, Denisovan ancestry today is depleted around functional regions of the genome and was therefore probably subject to a similar process of negative selection35.The third strongly supported admixture event in the ancestry of present-day people is from a second, distinct Denisovan population into the ancestors of present-day East Asian people, found in propor-tions in the order of 0.1%49,87,89,90. This population appears to have been more closely related to the Denisovans from Siberia87. East Asian popu-lations can thus trace very small amounts of ancestry to two distinct Denisovan-related groups.The frequency of archaic admixture outside of AfricaAnother strongly supported archaic admixture event comes from analyses of an approximately 40-thousand-year-old individual from Petera cu Oase in Romania91, who had a Neanderthal ancestor in the previous 4–6generations92. However, this Neanderthal admixture probably did not contribute to present-day ancestry92. Together with findings of admixture between Neanderthals and Denisovans61,93, these few, but direct, observations of admixture suggest no strong biologi-cal or behavioural barriers to admixture between modern humans, Neanderthals and Denisovans.Further archaic admixture events have been proposed, but lack consensus across the literature. These include additional pulses from Neanderthals46,71,73–77,86,94,95, Denisovans78,85,90 and unknown archaic ancestries96,97. An emerging notion, fuelled by the many hypothesized events in the literature, is that archaic admixture into modern human populations occurred frequently whenever these groups came into contact. Although such admixture complexity is certainly plausible, we argue that the currently well-supported events include only those described above.The current genomic data also do not support substantial unknown ‘ghost’ archaic ancestry in non-Africans89,98,99, for example from Homo erectus or other groups. More substantial amounts of such ancestry would give rise to an excess of ancestral variants in some non-African groups over others, detectable even without access to the source genome, a signal that can be confirmed for Denisovan ancestry10.Phase 2 and African originsThe second key phase is the diversification of present-day human ances-tries. 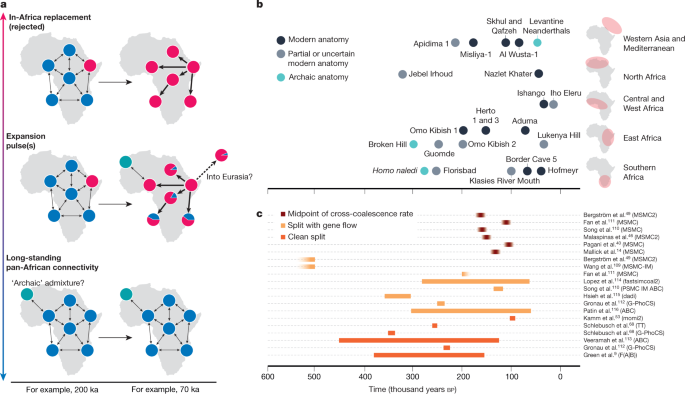 Although Africa was probably the centre of this process, neigh-bouring parts of Southwest Asia cannot be excluded as key areas of the history of the human population during the past few hundred thousand years100. However, an origin of present-day modern human diversity further away in Eurasia now seems to be highly unlikely. Here, we discuss what is known about modern human history in Africa between around 60 and 300ka in a conceptual framework of models (Fig.2a), among which only a complete replacement scenario inside Africa in the past 100ka can currently be excluded.The fossil record of modern human origins in AfricaFor the period from around 150 to 300ka, African fossil crania show great morphological diversity (Fig.2b). By contrast, Middle Stone Age technology, which emerged across Africa around 300ka, suggests similar patterns of behaviours across diverse human populations7,101. Specimens such as Jebel Irhoud 1 and 2 (Morocco, dated to around 315ka)7 and Omo Kibish 2 (Ethiopia, dated to around 195ka)8 are often placed on the modern human lineage, but lack a globular cranial vault. In fact, despite certain dental and mandibular traits pointing to H. sapiens affinity7, some studies place Irhoud 1 as closer to Neander-thals than to extant H. sapiens102,103. On the basis of the currently availa-ble evidence, a globular cranial vault appears only by about 150–200ka, when it is found in Omo Kibish 1 (Ethiopia, dated to around 195ka) and Herto 1 and 3 (Ethiopia, dated to around 160ka)8, although it might also be present in partial crania found at Apidima (Greece, dated to more than 210ka)21 and Guomde (Kenya, dated to around 240ka)8. The fragmentary Florisbad cranium (South Africa, dated to around 260ka, although this date requires confirmation104) is too incomplete to determine the extent of globularity, and its relationship to modern humans is uncertain105.With the scattered fossil evidence that is currently available, it is therefore unclear whether or not there was a specific evolutionary event between 200 and 300ka that originated a suite of ‘modern’ traits such as the globular cranial vault, bony chin on the mandible, and a narrower and less flared pelvis. One rationale has suggested that such traits were in place before the earliest separation of present-day ancestries. How-ever, if early separations were more gradual, as is increasingly likely and discussed further below, ‘modern’ traits could have become universal by later gene flow. The timing of modern human ancestry diversification will therefore provide only a weak constraint on the evolution of traits. |
|
|
|
Post by Admin on Aug 31, 2022 23:45:36 GMT
 The time depth of modern human population structureThe major distinguishable strands of present-day human ancestry could be summarized as including those associated with populations in West Africa, East Africa, the Central African rainforests, southern Africa and the world outside of Africa2,11,13,15,106,107. Many African populations can then be described as having ancestry from more than one of these strands, including non-African ancestry, reflecting complex admixture processes. The diversification of these African ancestries probably postdates the divergence of Neanderthal and Denisovan ancestors, as no differences in relatedness to archaic genomes between these lineages are apparent9,13. Understanding the diversification process of early modern humans more precisely has been a major focus in the study of human origins.One way to conceptualize the time depth of current human population structure is to focus on the earliest point in time at which groups existed that contributed more genetic ancestry to some present-day individuals than to others. There would also have been structure before this point, but any earlier groups would be symmetrically related to everyone alive today. The current absence of ancient DNA from Africa from before 15ka means that most insights into this question have come from estimates of divergence timing between present-day African groups, reliant on various modelling assumptions and therefore associated with consid-erable uncertainties. It is becoming clear that early divergences within Africa were not sudden splits, but were instead much more gradual, with long-standing gene flow over tens or even hundreds of thousands of years49,108,109. Estimates that can be interpreted as a midpoint of the separa-tion process result in relatively recent dates of 104 to 162ka14,40,46,49,110,111, whereas models assuming instantaneous separation without gene flow yield dates of 230–340ka9,68,112,113, and models that include gene flow yield dates of 125–340ka110–112,114–116 (Fig.2c). Different approaches may therefore partly capture different aspects of the gradual separation processes. We argue that it is not conceptually meaningful to describe the time depth of the population structure of early modern humans with point estimates, and that future studies should aim to be more explicit about what aspect of the separation process their estimates reflect.The question then becomes how we should describe the timing of the emergence of present-day human population structure. Looking backwards, the majority of the genetic ancestry of modern humans might converge between around 100 and 250ka, with a minority fraction of ancestries deriving from populations that had diverged earlier than that, possibly before 500ka49 or even one million years ago (Ma)109. Many different scenarios could underlie this observed timescale of shared ancestry and, other than rejecting models of recent African-wide replace-ment, current data do not clearly distinguish between them117,118 (Fig.2a).Possible ‘archaic’ admixture in AfricaQuestions about the time depth of the structure among present-day populations are inextricably linked to claims of admixture with more divergent human groups within Africa. Some of the few later fossils |
|
|
|
Post by Admin on Sept 3, 2022 0:34:51 GMT
in western and Central Africa (Iho Eleru, Nigeria, dated to around 13ka119,120, and Ishango, Democratic Republic of the Congo, dated to about 20–25ka121) show apparent archaic features, which may indicate either very late survival of early H. sapiens morphologies, or gene flow from archaic (with morphology outside the range observed today) human lineages.Several studies have suggested the presence of very deep population structure in Africa based on present-day genetic variation122–124, includ-ing suggestions of admixture from genetically unsampled ‘archaic’ human groups that are not closely related to Neanderthals or Deniso-vans115,125–128. In these genomic studies, the term ‘archaic’ is used not with reference to morphology as in the original meaning of the term, but rather to imply an early genetic divergence. The term ‘archaic’ is poten-tially problematic as it risks being misinterpreted as ‘less evolved’117. Given its long-standing usage, we argue that in a genomics context the term should only be applied to groups that are explicitly hypothesized to have separated chronologically from the majority of modern human ancestry at least as early as Neanderthals did.Most studies that aimed to identify highly divergent ancestry in Africa have looked for segments of the genome that are both unusu-ally long and deeply divergent from other segments122–124. Although these observations have been shown to match models of archaic admixture best in simulations115,125–128, it is challenging to rule out that such segments could represent the upper tails of the divergence distributions of high-diversity African populations. The concepts of ‘long-standing structure’ and ‘archaic admixture’ might thus be viewed as a continuum of models (Fig.2a). However, support for deep admix-ture also comes from derived alleles shared with Neanderthals at the rare and high-frequency ends of the spectrum in modern human popu-lations129. Some models of African population history have also included gene flow from lineages that diverged as early as, or before Neander-thals106, but simpler models have not been excluded. In our view, it is premature to refer to these variousfindings as archaic admixture, and they do not come with the same level of confidence as Neanderthal and Denisovan admixture, for which direct genomes from the source populations are available. Nonetheless, admixture from highly diver-gent groups inside Africa could help to explain the observed complex timing of early modern human separations.The search for a modern human birthplace.
We argue that, with current evidence, it is not possible to pinpoint more precisely where in Africa the common ancestors of present-day people lived. In the absence of a full timeseries that demonstrates how ancestry was distributed in the past, a strong line of evidence for an origin in a given geographical region could be if the majority of human ancestry was ‘nested’ inside the greater diversity of that region, accounting for admixture. However, although such a criterion currently identi-fies Africa as the birthplace of modern humans, it does not pinpoint a specific region inside Africa.Another rationale suggests that the highest levels of genetic diversity are found at the origin of expansions3,130,131, and this ‘serial founder’ model has been used to suggest a southern African origin of modern
|
|
|
|
Post by Admin on Sept 5, 2022 7:56:24 GMT
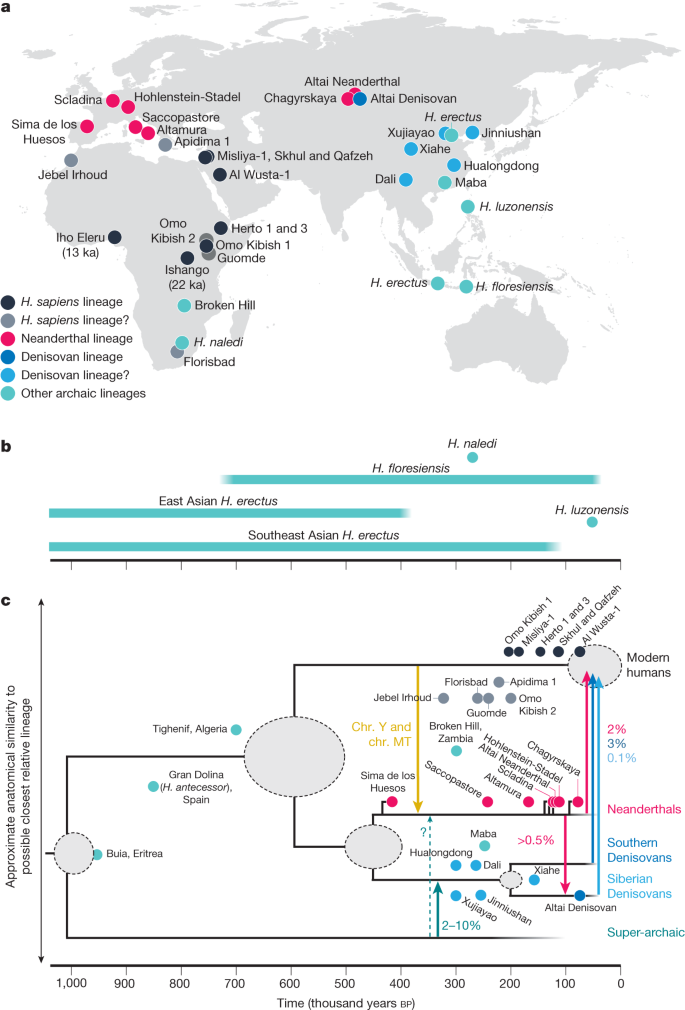
humans2,12. However, present-day diversity levels reflect not only loss of diversity due to population bottlenecks, but also increased diversity due to admixture132–134, as exemplified by how a pattern of greater diver-sity in Europe compared to East Asia today was not present in ancient populations58. Furthermore, diversity levels of major sub-Saharan populations are all within around 10% of each other, without strong geographical trends11,14,107,111. In a recent whole-genome panel14, the population with the highest diversity was the Central African Biaka, who have a history of recent admixture2,135.The tendency of populations from southern Africa to display the deepest divergence times, and to occupy the earliest branching posi-tions in tree-like models of history2,11,15,113, has also been interpreted as evidence for a southern African birthplace. However, trees are poor representations of genetic history, and branching events always have two symmetrical descendant branches, neither of which is more ances-tral than the other. More recent studies that allow for gene flow have suggested that strands of ancestry that are at least as divergent as southern African ancestry are present in western13 as well as Central and eastern Africa106. More importantly, as people are likely to have moved from where their ancestors lived more than 200ka, there is no strong expectation that the geographical location of the people carry-ing the most divergent ancestry today would correspond to a point of origin. Similarly, although a mitochondrial ‘Eve’, a hypothetical female ancestor of everyone alive today, will have existed and probably lived about 200ka1,136, the location where she, or her Y-chromosomal ‘Adam’ counterpart137, lived is not necessarily expected to be the birthplace of all human ancestry. Furthermore, the small mitochondrial history traces just one out of a multitude of paths through the greater human genealogy. In many other parts of the genome, the most divergent branch will be found elsewhere in Africa, or sometimes outside of Africa.For these reasons, current genomes simply do not contain enough information on where our early ancestors lived geographically. Recently, increasing attention has been given to the hypothesis that the ancestors of modern humans lived in distinct but interconnected populations across large parts of the African continent101,117, but such ‘pan-African’ origin hypotheses (Fig.2a) are similarly difficult to test against genomic evidence. A richer and geographically more repre-sentative fossil record, and ancient DNA or proteins from earlier time periods, might be more informative about past distributions of humans within Africa.A possible Late Pleistocene expansion across AfricaIn population history models that include deeply diverging ancestries in West13,106 and Central106 Africa, the second major ancestry in these regions tends to be related to East African populations. A specula-tive suggestion that could explain this is an expansion across Africa of an ancestry similar to that which expanded into Eurasia after around 60ka13,106. This could also explain the spread of the Y-chromosomal CT lineage, for which African and non-African lineages had a common ancestor around 65–80ka38,138. An analysis of divergence times between segments of present-day genomes similarly suggested substantial admixture from a source related to the ancestors of non-Africans, but lacking Neanderthal admixture, into all African populations studied139. This ancestry might never have left the African continent, but could represent an across-Africa expansion concurrent with the into-Eurasia expansion (Fig.2a), and its spread could potentially be a major contribu-tor to the complex genetic relationships observed among present-day African populations.Phase 1 and the divergence from archaic groupsThe most characteristic Neanderthal fossils are known from Europe from about 40 to 250ka140, with currently more limited time spans known across Asia, as far as southern Siberia61,141. Although the identity of Denisovans in the fossil record is currently poorly known, sediment mtDNA confirms their presence on the Tibetan plateau between at least around 60 and 100ka142, and the growing fossil record from China for the period from 200 to 600ka contains specimens that show differ-entiation from earlier Asian H. erectus143. The Dali skull has sometimes been grouped with Homo heidelbergensis, but shows a combination of a massive supraorbital torus, an archaic-shaped cranial vault and a rather modern-looking, but very wide face144. This is a distinctive morph, which is perhaps mirrored in other Chinese fossil crania such as those from Jinniushan145 and Hualongdong146 that also lack characteristic features of H. erectus, H. heidelbergensis and Neanderthals. These could thus represent candidates for early Denisovans, along with the mandibles from Xiahe and Penghu147. Neanderthals and Denisovans share some ancestry after the separation from modern humans, but they probably diverged from each other before 400ka61,148.
Analyses of genomes have identified, in addition to Neanderthal, Denisovan and modern human ancestry, a fourth, highly distinct ances-try that would have existed during the time period before 300ka (Fig.3), but which currently cannot be linked to any populations apparent in the fossil record: the ‘super-archaic’ ancestry proposed to be pre-sent in Denisovan genomes71. The primary evidence is that all modern humans, including African populations with little or no Neanderthal admixture, share more genetic variants with Neanderthals than with Denisovans71, and particularly so for variants that became fixed in early modern humans—with super-archaic ancestry diluting the frequency of these variants in Denisovans. This super-archaic group would have diverged from the common ancestor of modern humans, Neanderthals and Denisovans between 0.9 and 1.4Ma61,149, or earlier61,150. Denisovans also carried a highly distinct mitochondrial lineage that diverged from others around 0.7–1.4Ma151,152 and that was probably obtained from this population. Although it is tempting to speculate that this super-archaic population could correspond to H. erectus or some related group, its genetic divergence seems too recent to align with the first appearance of fossil H. erectus at least around 1.8Ma153. Populations related to Homo antecessor154 might be an alternative.The ancestors of Neanderthals and Denisovans are estimated to have diverged from the ancestors of modern humans between 500 and 700ka9,49,53,61,62,68,155. Although it has been suggested that this was a sudden rather than agradual separation process49, evidence against complete genetic separation since more than 500ka comes from the mtDNA of modern humans and Neanderthals diverging only around 350–450ka151,152,156,157, and a similar timeframe for Y chromosomes158. This apparent discrepancy could be explained if the uniparental chro-mosomes moved between the ancestors of Neanderthals and modern humans by gene flow, in either direction, at some point after about 450ka.An important data point in resolving this history is DNA from the more than 400-thousand-year-old159 site of Sima de los Huesos in Spain, where skeletons with Neanderthal-like physical traits also have an affin-ity to Neanderthal rather than Denisovan genomes148. However, indi-viduals from Sima de los Huesos carry a version of the highly diverged mtDNA lineage that is found in Denisovans156, suggesting that it once could have been carried by all early Neanderthals, but was replaced by gene flow from the ancestors of modern humans148,152,158. Later Nean-derthal mtDNA lineages diversified around 270ka, suggesting that the gene flow into Neanderthals occurred before this date152, although it is possible that part of this diversity was present earlier.Gene flow from the modern human lineage into Siberian, but not European, Neanderthals has been suggested160, but higher-quality genomes have not corroborated this62. Instead, later studies have sta-tistically inferred modern human gene flow on the order of a few per cent into the ancestors of all Neanderthals studied to date63,160,161. Such inferences are subject to many of the same modelling challenges as analyses of archaic admixture in Africa. Regardless, any such gene flow would need to have come from a population that diverged mostly before the diversification of present-day modern human ancestries, more than 200ka, but could correspond to the same event that led to the transfer of the uniparental chromosomes.Three possible ancestry strands that contributed to Neanderthals and Denisovans have therefore been hypothesized, with the strands having different degrees of divergence from present-day humans: (1) ‘super-archaic’ ancestry that diverged around 1Ma; (2) the originally hypothesized ‘middle’ archaic ancestry, which diverged from the ances-tors of modern humans between around 500–700ka to give rise to both Denisovan and Neanderthal ancestry; and (3) ‘recent gene flow from modern human ancestors’ around 200–400ka. Super-archaic ancestry has been inferred for Denisovans61, and recent gene flow for Neanderthals148, but it is also possible that both archaic populations carried both ofthese strands of ancestry in different proportions88. Although it is commonly believed that both Denisovans and Neander-thals derived the majority of their ancestry from the ‘middle’ archaic population, it may be possible that the inferred archaic–modern human divergence of 500–700ka49,53,61,62,68 is due to statistical averaging of the ‘super-archaic’ and ‘recent gene flow’ ancestries. No ‘middle’ popula-tion and expansion of Neanderthal ancestors between 500 and 700ka would be necessary in that alternative scenario88.
|
|
|
|
Post by Admin on Sept 6, 2022 9:56:25 GMT
The last common ancestor of modern and archaic humansOnce the majority of the ancestry of modern humans, Neanderthals and Denisovans has converged before 500ka, genomes provide little or no information about who those common ancestors were. Fossils from around 300–700ka have revealed many anatomically distinct groups of humans, and the period has been called the ‘muddle in the middle’162 of human evolution. It is impossible to identify any early Middle Pleistocene fossils as definitively representing the common ancestral population for H. sapiens, Neanderthals and Denisovans, but it is possible to identify groups that probably are not, namely Asian H. erectus, facially derived H. heidelbergensis across Africa and West Eurasia, and the Neanderthal-like Sima de los Huesos hominins. Possible alternative candidates for our early ancestors might include H. antecessor from Europe, the Tighenif fossils from Northwest Africa163 and the Buia material from Northeast Africa164.Although it is commonly assumed that our ancestors would have lived in Africa before 500ka, it is still too soon to exclude that they could have lived in Eurasia. A Eurasian origin during this period would also require fewer migrations between Africa and Eurasia to explain currently understood relationships between modern human, Nean-derthal, Denisovan and the super-archaic ancestries88. Proteomic data from European H. antecessor165, which shows the potential of ancient protein preservation in the deep past, suggests that it might have been closely related to the common ancestor, but the ancestry information provided by dental enamel proteins is still of low resolution. In any case, with the earliest generally accepted evidence of hominins outside of Africa at around 2Ma166, the fossil record strongly suggests that all human ancestors before this point, until the common ancestor with chimpanzees, lived in Africa.OutlookAlthough a common understanding views modern humans as having an African origin in the last few hundred thousand years ago (for example, ‘200 ka’), what such an ‘origin’ entails is often not well defined. It is increas-ingly important to differentiate the evolution of traits—that is, when our ancestors became sufficiently similar to present-day humans in terms of anatomy6–8,167, behaviour, physiology or cognition—from genetic ancestry. Definitions from a genetic perspective may instead focus on a period in time in which most of the genetic ancestry of present-day people was found in a specific geographical area, with or without a particular set of traits. Asking when and where modern human ancestry originated is a different question from asking when and where modern humans, as defined through our traits, originated, and the answers to the first ques-tion that we have reviewed here may only weakly inform on the latter. Any strict definition of origin thus risks oversimplifying the continuous and complex, and in many aspects unknown, nature of the deep human past. For example, current evidence identifies Africa and Southwest Asia as the region of origin of humans during the period of 100–300ka, but does not yet provide further geographical precision, and before 300ka there is even greater uncertainty about where our ancestors lived.Over the next decade, these insights will probably also shift the geographical focus of palaeoanthropological fieldwork to regions that previously have been considered peripheral to perceived cen-tres of human evolution, such as Central and West Africa, the Indian subcontinent and Southeast Asia. As more spatially and temporally representative palaeoanthropological and genetic data from across Africa and the rest of the world become available, it will be possible to refine our understanding of ancestry through the human past as described here. The success of direct genetic analysis so far highlights the importance of a wider ancient genetic record. This will require continued technological improvements in ancient DNA retrieval from skeletal material60,71,168, biomolecular screening of fragmentary assem-blages for human material93,169, analysis of sedimentary DNA142,170 and improvements in the evolutionary information provided by ancient proteins147,165. Interdisciplinary analysis of this combined record will undoubtedly reveal new surprises about the roots of modern human ancestry.
|
|

