|
|
Post by Admin on Nov 13, 2022 0:30:57 GMT
Fig. 2: Relative eukaryote abundance across all samples using SSU, LSU and SSU + LSU reference databases. 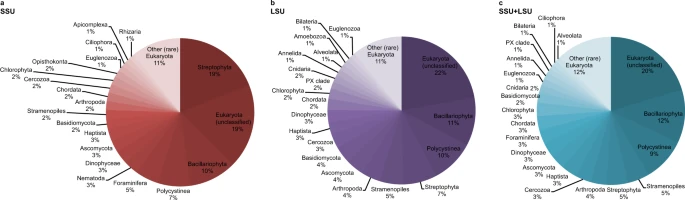 Pie charts were generated based on relative abundances (phylum level) determined after running shotgun data against the SILVA 132 SSU (a), LSU (b), and combined SILVA SSU + LSU (c) databases. Taxa that contributed <1% on average across all samples are summarised as’Other (rare) Eukaryota’. Source data are provided as a Source Data file. sedaDNA-derived eukaryote composition at the three sites At each site, we recovered very few eukaryote reads (0–2) from the mudline samples, maximum number of reads from intermediate depth samples (670 reads at U1534 5.46 mbsf, 7302 reads at U1536 4.46 mbsf, and 22,631 reads at U1538 12.05 mbsf), and few reads in the bottom-most samples (37 reads at U1534, 52 reads in our oldest sample from U1536, and 94 reads from our oldest sample at U1538, dated to ~0.7–2.5 Ma, 1 Ma, and 417 ka, respectively (for details on age determination see Methods section). Across all sites, Eukaryota (category not further classified) were the most abundant (~20% on average across all samples), followed by Bacillariophyta (diatoms, ~11.7%), Polycystinea (which include the Radiolarians, 9.5%), Stramenopiles (not further classified, 4.8%), Streptophyta (4.6%), Arthropoda (3.5%), Cercozoa (3.4%), Haptista (3.4%), Ascomycota (3.25%), Dinophyceae (3.2%), Foraminifera (3.1%), Chordata (3%), Chlorophyta (3%), Basidiomycota (2.3%), Cnidaria (1.5%), Euglenozoa (1.5%), Annelida (1.5%), PX clade (1.1%), Bilateria (1%), Ciliophora (1%) and Alveolata (1%) (Fig. 2). All other phyla contributed <1% on average across all samples to the eukaryote composition (Fig. 2). For details on taxonomy used and assignments see Methods section. Using the SSU + LSU alignments, we identified eukaryotes in all but three samples, and found that eukaryote composition changed with depth at all three sites (Fig. 3). At all sites, Eukaryota (not further classified) contributed most of the assigned reads, followed by diatoms (Bacillariophyta). At Site U1538, the relative abundance of diatoms increased from <10% in sediments as old as 34 ka to ~50% of all eukaryotes in samples younger than 12.7 ka (with ~33% around 14.5 ka; Fig. 3c). The relative abundance of diatoms also increases in younger sediment at Site U1536, however, lower sampling resolution only allows the timing of this shift to be constrained between 53 and 4 ka (Fig. 3b). No obvious increase in the relative abundance of diatoms was observed using sedaDNA at Site U1534 (Fig. 3a). While most of our sedaDNA samples were younger than 540 ka, we examined the two samples with an estimated age of ~0.7–2.5 Ma (Site U1534) and ~1 Ma (U1536) and acquired few reads (37 and 52, respectively) and few taxa, primarily consisting of Fungi (Asco-, Basidio-, Chytridio- and Mucoromycota), unclassified Eukaryota, and a few rarer groups (e.g., Chordata, Metazoa, Cercozoa, PX clade, Streptophyta and Chlorophyta; Fig. 3a, b and Source Data File). |
|
|
|
Post by Admin on Nov 24, 2022 21:01:01 GMT
Figure 3 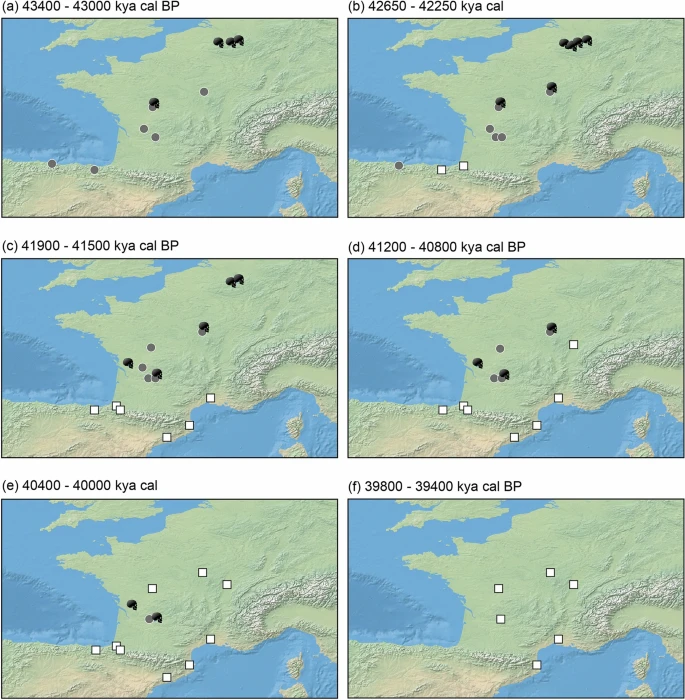 Geographic appearance of dated occurrences for the Châtelperronian (grey circles), Protoaurignacian (white squares), and directly-dated Neandertals (black skulls) in the study region between 43,400 (a) and 39,400 (f) years cal BP. Figure was produced using the ‘spatio-temporal modeller’ function in the QGIS software (v4.4) (QGIS Geographic Information System, QGIS Association, 2021) and compiled in Adobe Illustrator. Using OLE modelling to infer the ‘origination’ time of the Protoaurignacian and the ‘extinction’ time of the Châtelperronian and Neandertals in the region We had three objectives, with each requiring its own OLE model and respective sample (see ‘Methods’): 1. Estimating the origination date of the Protoaurignacian in France and northern Spain. The nine oldest Protoaurignacian dates from nine discrete archaeological sites are entered into this OLE model (Supplementary Table S2, Supplementary Fig. S7) which is run in the reverse temporal direction. 2. Estimating the end date of the Châtelperronian in France and northern Spain. The eight youngest Châtelperronian dates from seven discrete archaeological sites are entered into this OLE model (Supplementary Table S3, Supplementary Fig. S6) which is run in the forward temporal direction. 3. Estimating the extinction date of regional Neandertals. Ten direct dates of late Neandertal individuals from France (n = 4) and Belgium (n = 6) are entered into this OLE model which is run in the forward temporal direction (Supplementary Table S4, Supplementary Fig. S5). OLE modelling infers the Protoaurignacian to have likely emerged in France and northern Spain by 42,269 to 42,653 years cal BP. The upper bound of this TO date range is defined by the resampling technique, while the lower uses the central tendency (mean) dates derived from the C14 date range. As explained earlier, we consider the resampling estimates to better account for the range uncertainly inherent to C14 dating. TCI dates, beyond which the Protoaurignacian only has a 5% chance of preceding this point, provide a bracket of 43,394–44,172 years cal BP. Upper and lower bounds were again defined by the resampling technique and central tendency dates (respectively). OLE modelling estimates the Châtelperronian to have disappeared by 39,894 to 39,798 years cal BP. The upper bound of this TO date range is defined by the resampling technique, while the lower uses the central tendency (mean) dates. TCI dates, beyond which the Châtelperronian only has a 5% chance of following this point, provide a bracket of 37,838–37,572 years cal BP. Again, upper and lower bounds were defined by the resampling and central tendency dates respectively. OLE modelling infers the localised extinction of Neandertals in France and Belgium to have occurred between 40,870 to 40,457 years cal BP. The upper bound of this TE date range is defined by the resampling technique, while the lower uses the central tendency (mean) dates. TCI dates, beyond which Neandertals only have a 5% chance of following this point, provide a bracket of 39,688 to 38,752 years cal BP—with the upper and lower bounds again defined by the resampling and central tendency techniques, respectively. Across all OLE models, the resampling approach extended temporal ranges by several hundred years compared to the central tendency (mean) based estimates. The results of the 10,000 resampling iterations for each model are illustrated in Fig. 4 and the raw data is available in full (Supplementary Data S2). Combined, OLE modelling suggests the Protoaurignacian to have emerged around 1399–2196 and 2375–2855 years before Neandertals and the Châtelperronian industry (respectively) disappeared from the region. |
|
|
|
Post by Admin on Nov 28, 2022 21:48:45 GMT
Figure 4 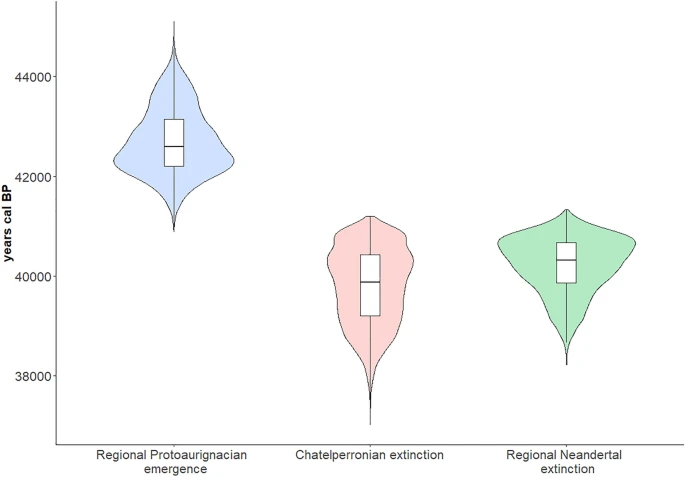 Resampling results for the three OLE models: modelled Protoaurignacian regional emergence time (left), modelled Châtelperronian extinction time (centre), and modelled Neandertal regional extinction time (right). The horizontal bar in each respective boxplot represents the mean value of the 10,000 resampling iterations referred to in the text. Figure was produced using the ‘ggplot2’ package in R (version 4.0.3). Discussion Based on OLE modelling of their respective ‘origination’ and ‘extinction’ dates, the Protoaurignacian potentially appeared around 1400–2900 years before Neandertals and the Châtelperronian industry disappeared from France and northern Spain. This is largely consistent with previous estimates, and reaffirms the duration of co-existence between these groups during the early western European Upper Palaeolithic. Furthermore, it may indicate that our understanding of the timing of these occupations may not be suffering from significant gaps in the archaeological record. The oldest calibrated age ranges from well-dated Protoaurignacian assemblages initially form a cluster at the southern and northern limits of France and Spain respectively, overlapping with dates produced for Châtelperronian assemblages in the central-northern parts of France. This may indicate that the chronological overlap may have been geographically structured, with the Protoaurignacian following a south to north pattern of appearance. Moreover, based on OLE estimates produced using directly-dated Neandertal fossil remains, the onset of the regional Protoaurignacian is modelled to have preceded the extinction of Neandertals in this region by upwards of 2200 years. These results are perhaps not surprising given the nature of probability ranges for calibrated radiocarbon determinations produced for this period—which is temporally situated near the upper acceptable limit of radiocarbon dating (circa 50 kya)48. However, the fact that the OLE ‘extinction’ and ‘origination’ estimates produced here do not go far beyond the ranges identified in the calibrated radiocarbon dates themselves is notable, and is likely related to the temporal spacings observed for each of the occurrences. In each case the latest/oldest series of dates for each category (Protoaurignacian, Châtelperronian, Neandertal) reflect a narrow temporal band with little variation and inter-date spacing (ie., the dates are chronologically close), after which no additional dated occurrences are known. This has two potential implications. Firstly, that the oldest and/or youngest known dates for each industry are likely close to the true origination and/or extinction date of that industry34. Secondly, and relatedly, the true emergence and/or extinction dates may in some cases be slightly more conservative than the upper limits of the oldest and/or youngest calibrated dates themselves. This is perhaps particularly relevant for the Châtelperronian, which is widely acknowledged as largely reflecting relatively ephemeral and short-lived occupations15—with the exception of some notable examples16,38. Of course, there are limitations in this analysis which require consideration. The most obvious concerns the sample size of archaeological sites included in this work—which was militated by the decision to employ strict, conservative sampling requirements for the radiocarbon datasets (see ‘Methods’). And while we acknowledge that the sample considered here reflects a sample of known Châtelperronian and Protoaurignacian occurrences within this region, it does cover their known geographic distribution. Moreover, OLE works best with limited datasets, such as this. A second potential limitation concerns the radiocarbon determinations themselves. Any model is, of course, only as reliable as the data entered into it. The assumption taken here is that the age ranges entered into the models reflect meaningful datapoints for the chronological presence of these occurrences. This, in time, may change as the duration of these industries is increasingly refined. At present however there is no clear evidence to doubt the reliability of the radiocarbon determinations used in this study, but future work may necessitate the revision of this model as more sites are dated—or re-dated—and further methodological advancements are made. From an archaeological perspective, of relevance to these results is the acknowledged presence of bladelet technologies, osseus artefacts, and personal ornaments within a growing number of Châtelperronian and Protoaurignacian contexts. Unanimously seen as a trademark of the Protoaurignacian techno-complex (with the laterally retouched Dufour bladelet [sub-type Dufour] fossile directeur commonly constituting a substantial portion of Protoaurignacian assemblages), evidence for some form of intentional bladelet production and/or modification within the Châtelperronian has now been reported from at least five open-air sites14,17,49,50 and six cave sites16,42,49,51,52,53. To what extent (if any) these similarities represent some form of connection between these industries remains unclear, but the potential contemporaneity of the groups producing these assemblages is certainly of relevance. Of course, the results presented here do not aid in answering the question of which human group(s) were responsible for producing these industries, but the temporal and geographic proximity of directly-dated Neandertal remains to both Châtelperronian and AMH-attributed Protoaurignacian assemblages in the region is—in the current state of knowledge—difficult to overlook. With this being said, the recent chronological re-evaluation of late-dating Belgian Neandertals has convincingly demonstrated that they are likely substantially older than previously thought (Fig. 5)54. With this development, the Neandertals from France included in this study are now among the latest directly-dated Neandertals identified throughout the inferred geographic distribution of this human group. This raises an important consideration: it is possible that future work employing emerging radiocarbon dating techniques designed to further mitigate anthropogenic and/or natural contamination issues (e.g. Compound Specific Radiocarbon Analysis) may, in time, either confirm or revise their currently accepted ages. |
|
|
|
Post by Admin on Dec 4, 2022 21:54:09 GMT
Figure 5 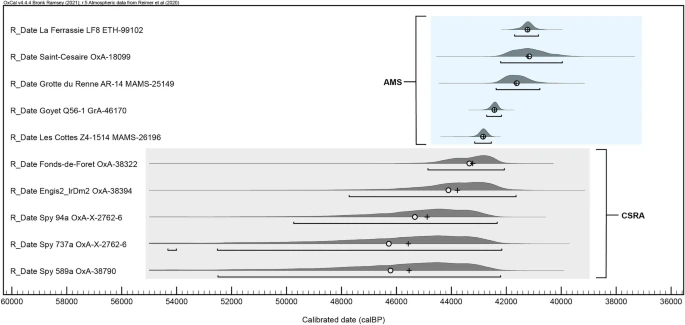 Regardless, considering the rapidly evolving understanding of the European demographic landscape preceding the onset of the Aurignacian techno-complex sensu lato3,4,5,7, it is clear that more work is needed to evaluate the biological identity and the cultural connections, if any, between the makers of archaeological industries across the European landmass during this period. However, at present, the only hominin species to as of yet be associated with Châtelperronian assemblages, based on both morphological and genetic evidence, is Neandertals. For the Protoaurignacian, the case is reversed—with the only published hominin association in a Protoaurignacian context being two deciduous teeth from Riparo Bombrini and Grotta di Fumane caves in Italy which have been attributed as Homo sapiens based on morphological criteria and mitochondrial DNA, respectively55. With this said, at present, the reality is that most Protoaurignacian assemblages are serving simply as well-accepted proxies for the presence of Homo sapiens—but the validity of this unilateral association is, in the current state of evidence, far from certain. In fact, in many ways the same can be said for Châtelperronian assemblages and their unilateral association with Neandertals13,24,53. With specific reference to the ongoing proliferation of paleogenetic research (including sedimentary aDNA analysis) and the increase in use of Zooarchaeology by Mass Spectrometry (ZooMS), future work will undoubtedly shed new light on the biological makers of these industries. The onset of the Châtelperronian, in the current state of knowledge, appears to clearly pre-date the appearance of the Protoaurignacian—both regionally and at a European scale. However, the spatio-temporal overlap of these assemblages in France and northern Spain—and their potential overlap with multiple directly-dated Neandertals from the region—lend credence to the idea that the early stages of the Upper Palaeolithic of this region may have involved the proximal co-existence of different human groups, likely irrespective of their biological classification. Conclusion Optimal linear estimation modelling predicts the appearance of Homo sapiens and the Protoaurignacian in France and northern Spain at 42,269 to 42,653 years cal BP, and the ‘extinction’ of the Châtelperronian and regional Neandertals at 39,894 to 39,798 and 40,870 to 40,457 respectively—suggesting a possible overlap of around 1400 to 2800 years between these human groups in the region. This is consistent with Bayesian estimates, and independently reaffirms the duration of co-existence between these groups during the initial Upper Palaeolithic of this region—indicating that our understanding of the timing of these occupations may not be suffering from significant gaps in the record. Additionally, this chronological overlap appears to be geographically structured, with the Protoaurignacian following a south to north pattern of appearance. Taken together, these observations strengthen the proposition that the initial Upper Palaeolithic in this region likely involved a period of co-existence between Neandertals and Homo sapiens. The precise nature of this co-existence, however, remains to be resolved. |
|
|
|
Post by Admin on Dec 7, 2022 20:54:04 GMT
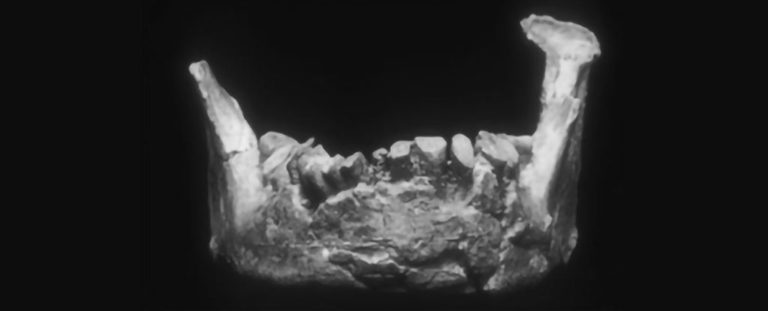 An ancient jawbone previously thought to have belonged to a Neanderthal may force a rethink on the history of modern humans in Europe. A new analysis of the broken mandible reveals that it has nothing in common with other Neanderthal remains. Rather, it could belong to a Homo sapiens – and, since it's dated to between 45,000 to 66,000 years ago, might be the oldest known piece of our species' anatomy on the European continent. The bone itself was found in 1887 in the town of Banyoles in Spain, for which it is nicknamed. Since then, scientists have studied it pretty extensively, dating it to a timeframe in the Late Pleistocene when the region that is now Europe was predominantly populated by Neanderthals (Homo neanderthalensis). This, and the archaic shape of the bone, led scientists to the conclusion that Banyoles in fact belonged to a Neanderthal. "The mandible has been studied throughout the past century and was long considered to be a Neandertal based on its age and location, and the fact that it lacks one of the diagnostic features of Homo sapiens: a chin," says palaeoanthropologist Brian Keeling of Binghamton University in the US. 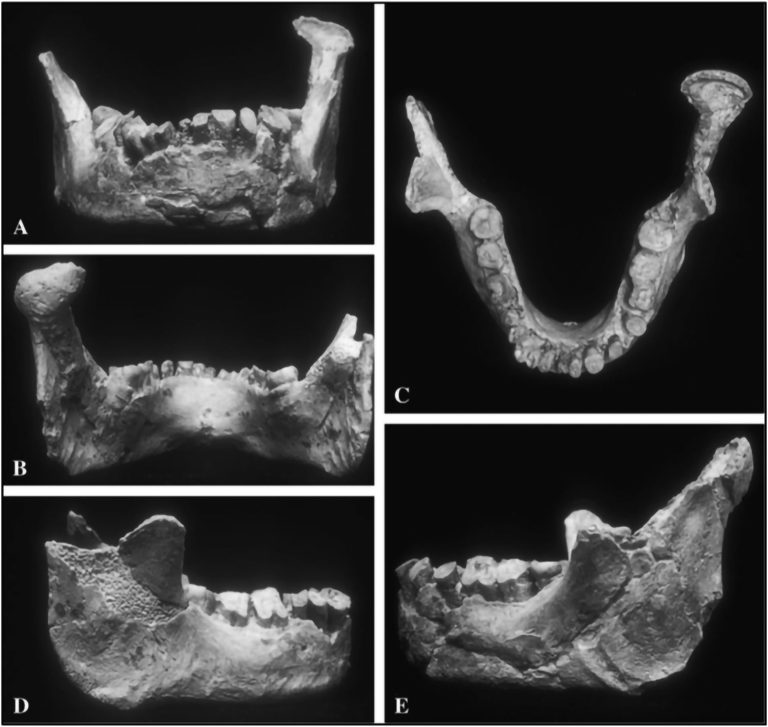 Keeling and his colleagues undertook a thorough investigation of the bone using a process called three-dimensional geometric morphometric analysis. This is a non-invasive protocol that involves going over the shape of a bone in exhaustive detail, mapping its features and comparing them to other remains. They took high-resolution 3D scans, and used these not just to study the bone, but to reconstruct the missing pieces. Then they compared Banyoles to the mandibles of Neanderthals and modern humans. "Our results found something quite surprising," Keeling says. "Banyoles shared no distinct Neanderthal traits and did not overlap with Neanderthals in its overall shape." It seemed more consistent with the jawbones of our own branch of the family tree, except for one detail: the absent chin. Since a chin is considered a defining feature of Homo sapiens compared to other archaic humans, this presented a problem. In addition, Banyoles also shared features with ancient hominins that inhabited Europe hundreds of thousands of years ago. The researchers compared the bone to one from an early modern human from around 37,000 to 42,000 years ago whose remains were found in Romania. It's known for having Neanderthal features, but also has a chin. DNA analysis of that jawbone showed the DNA included sequences from a single Neanderthal ancestor who lived four or six generations prior – which likely explains its mixed features. Since Banyoles doesn't have Neanderthal features, the team concluded that its strange shape is unlikely to be because the individual was a hybrid. Comparison with earlier Homo sapiens bones from Africa showed that these individuals had less pronounced chins than we do now. So there are two possibilities. Either Banyoles was a Homo sapiens from a previously unknown group that coexisted with Neanderthals in Late Pleistocene Europe. Or it was a hybrid between Homo sapiens of this unknown group and a yet-to-be-identified ancient human. Only one thing is known for sure: that Banyoles was not a Neanderthal. There is one way to resolve the mystery, the researchers say – try to extract some DNA from the bone or one of the teeth, and sequence it. "If Banyoles is really a member of our species, this prehistoric human would represent the earliest Homo sapiens ever documented in Europe," Keeling says. The research has been published in the Journal of Human Evolution. Reassessment of the human mandible from Banyoles (Girona, Spain) www.sciencedirect.com/science/article/abs/pii/S0047248422001518 |
|

