|
|
Post by Admin on Jun 7, 2022 19:21:26 GMT
Homo Erectus: The First Homo Sapiens? 9,980 views Jun 7, 2022 For most of the 20th century, much of the western academic world believed in the Out of Africa theory of human evolution. About 2 million years ago Homo erectus left Africa and spread throughout Europe and Asia, becoming the first alien hominid. Homo erectus evolved into the Neanderthals and Denisovans, while in Asia most Peking Man stopped evolving, with the exception of Dragon Man. According to paleoanthropologist Chris Stringer, most Asian Homo erectus stopped evolving - with the exception of a small group in the Indonesian archipelago that branched off to become Homo floresiensis, and the Homo erectus that stayed in Africa continued to evolve and eventually became Homo sapiens and Homo rhodosiensis.  The reversal of human phylogeny: Homo left Africa as erectus, came back as sapiens sapiens The reversal of human phylogeny: Homo left Africa as erectus, came back as sapiens sapiensAbstract Background The molecular out of Africa hypothesis, OOAH, has been considered as an established fact amid population geneticists for some 25–30 years despite the early concern with it among phylogeneticists with experience beyond that of Homo. The palaeontological support for the hypothesis is also questionable, a circumstance that in the light of expanding Eurasian palaeontological knowledge has become accentuated through the last decades. Results The direction of evolution in the phylogenetic tree of modern humans (Homo sapiens sapiens, Hss) was established inter alia by applying progressive phylogenetic analysis to an mtDNA sampling that included a Eurasian, Lund, and the African Mbuti, San and Yoruba. The examination identified the African populations as paraphyletic, thereby compromising the OOAH. The finding, which was consistent with the out of Eurasia hypothesis, OOEH, was corroborated by the mtDNA introgression from Hss into Hsnn (Neanderthals) that demonstrated the temporal and physical Eurasian coexistence of the two lineages. The results are consistent with the palaeontologically established presence of H. erectus in Eurasia, a Eurasian divergence between H. sapiens and H. antecessor ≈ 850,000 YBP, an Hs divergence between Hss and Hsn (Neanderthals + Denisovans) ≈ 800,000 YBP, an mtDNA introgression from Hss into Hsnn* ≈ 500,000 YBP and an Eurasian divergence among the ancestors of extant Hss ≈ 250,000 YBP at the exodus of Mbuti/San into Africa. Conclusions The present study showed that Eurasia was not the receiver but the donor in Hss evolution. The findings that Homo left Africa as erectus and returned as sapiens sapiens constitute a change in the understanding of Hs evolution to one that conforms to the extensive Eurasian record of Hs palaeontology and archaeology. |
|
|
|
Post by Admin on Jun 7, 2022 20:11:52 GMT
Background
Two molecular studies [1, 2] that came from the same laboratory were highly instrumental in the promotion of the out of Africa hypothesis, OOAH, ≈ 30 years ago. The hypothesis became gradually a mainstay in the discussion of human evolution and dispersal in spite of the absence of palaeontological support for it and the circumstance that reanalysis of the data [3,4,5] did not favour the phylogenies that constituted the foundation for the OOAH.
The OOAH was addressed previously in two studies [6, 7] in which the author maintained on the basis of molecular and palaeontological data that recent humans, Hss, had originated in Eurasia (the Out of Eurasia ypothesis, OOEH) and dispersed from there to other parts of the world including Africa. In the present study the two hypotheses were compared in the light of the identity of the basal divergence of the Hss tree as established in genomic analyses [8, 9]. One of these studies [8, Fig. 2a; see also 7, Table 1] identified this divergence as falling between all non-Africans and the African branch of Mbuti/San. The results of the other study [9], which included the Baka population as a representative of basal African lineages and Koinabe (Sahul) as a representative of non-Africans, complied with that finding. The implications of the results were detailed in a previous examination of OOAH and OOEH [7]. As concluded there the reorientation of the Hss tree was inconsistent with the understanding that the ancestors of non-Africans had originated by an exodus out of Africa.
The steady progress in the molecular analyses of Hss and Hsn (Hsnn + Hsnd) [10,11,12,13,14,15,16,17] has allowed an extended examination of OOAH and OOEH. With the palaeontology of Hsn (Hsnn + Hsnd) strictly limited to Eurasia [18,19,20,21] the molecular exchanges between Hss and Hsn are consistent with a continuous existence in Eurasia of an Hss population that is much older than the basal divergence among extant Hss populations, i.e. that between non-Africans and Mbuti/San. The palaeontological support for OOEH is upheld similarly by the findings of Eurasian Hss fossils [e.g. 22] whose ages, ≈ 200,000 years, surpass by 130,000–140,000 years the age that is commonly associated with the dispersal of Hss out of Africa. These finds and the Eurasian placement of the root of the Hss tree constitute a natural extension of the extensive Eurasian, palaeontological and archaeological records that relate to the origin and evolution of Hss and Hsn [23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72]. The existence and hence the phylogenetic implications of these studies have remained disregarded, however, among the proponents of OOAH.
Here the Out of Africa and the Out of Eurasia hypotheses were compared inter alia in the light of a new approach, progressive phylogenetic analysis (PPA), that allowed the examination of the mtDNA relationship among the African Hss populations in the light of new palaeontological and molecular findings. The particular result of this examination was that it disrupted the monophyly of the African populations, therewith compromising the cornerstone of OOAH.
|
|
|
|
Post by Admin on Jun 8, 2022 19:29:42 GMT
Results and discussion The OOEH phylogeny of basal Hss relationships The essence of the nuDNA phylogeny behind OOEH and the evolution of Hs (Hss + Hsn) is shown in Fig. 1 with the Eurasian lineages marked blue and the African red. The phylogeny encompasses an African exodus of H. erectus > 2 MYBP and a continuous evolution of H. erectus in Eurasia leading to a population that diverged into H. antecessor [66, 73] and H. sapiens ≈ 850,000 YBP. The arrowheads that lead from Hss into the Hsnn* branch in Fig. 1 have a particular phylogenetic significance for the interpretation of Hs evolution as they signify the mtDNA introgression that took place from Hss into Hsnn* ≈ 500,000 YBP, a transfer that could only come about in agreement with the physical coexistence of Hss and Hsnn*. In the light of the limitation of Hsnn to Eurasia the introgression and the continuous evolution of both Hss and Hsn can only be placed in this continent in accordance with OOEH. Fig. 1 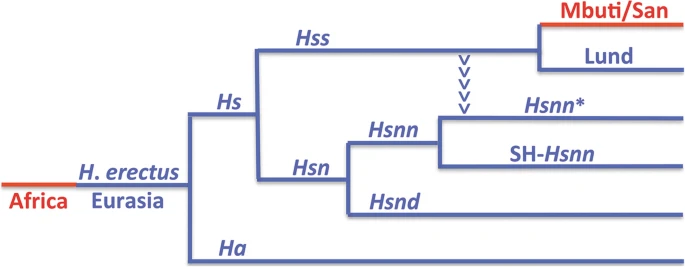 The nuDNA relationship of Hs, Homo sapiens, and Ha, Homo antecessor, with Eurasian lineages in blue and African in red. Hss (H. s. sapiens); Hsn (H. s. neanderthalensis); Hsnn (H. s. n. neanderthalensis, Neanderthals proper); Hsnd (H. s. n. denisova, Denisovans). The divergence between Hs and Ha has been dated to ≈ 850,000 YBP [73] and that between Hss and Hsn to ≈ 800,000 YBP. The arrowheads mark the mtDNA introgression that took place from Hss into Hsnn* ≈ 500,000 YBP, i.e. more recently than the Hsnn divergence into Hsnn* and SH-Hsnn. The basal divergence among recent Hss, as represented by the Eurasian Lund and the African Mbuti/San is placed in Eurasia at ≈ 250,000 YBP Figure 2 summarizes the relationships connected to the evolution of Hs (Hss + Hsn) as established in analysis of complete mtDNAs. Tree 2a demonstrates the basal phylogeny of the sampling, while the trees 2b–d show only the Hss part of the complete Hs tree. The analysis was restricted to the minimum number of taxa required for the examination: a chimpanzee, Pan troglodytes [74] for rooting the tree, a Neanderthal proper, Hsnn [75], a Denisovan, Hsnd [76], Mbuti, San and a series of Yoruba (all African) and Lund, the first described non-chimaeric human mtDNA sequence [74], as a representative of non-Africans. The inclusion of the Yoruba individuals rested upon the circumstance that Yoruba is commonly taken as the African ancestor of non-African populations in studies that acknowledge OOAH [e.g. 77]. |
|
|
|
Post by Admin on Jun 9, 2022 18:02:58 GMT
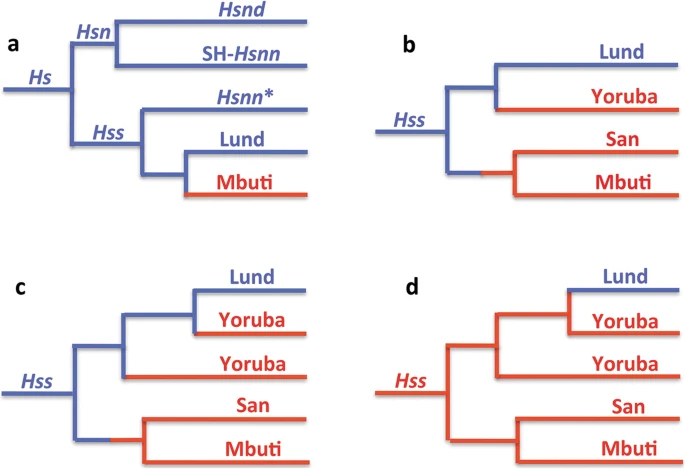 Figure 2 The essence of Hs mtDNA relationships as resolved in PPA (Progressive phylogenetic analysis) with Eurasian lineages in blue and African in red. a: The Hs tree, rooted with the chimpanzee, shows a basal divergence between Hsn and Hss with Hsnd and SH-Hsnn on a common branch and Hsnn* joining the Hss branch as the result of the mtDNA introgression from Hss into Hsnn* ≈ 500,000 YBP. Mbuti and Lund make up the basal divergence among extant Hss. b and c: The inclusion of San and the Yorubas joined San on a common branch with Mbuti, whereas the Yorubas joined the Lund branch at separate positions in accordance with the paraphyly of the African populations. d: The OOAH tree with Hss origin in Africa followed by a late exodus into Eurasia. The OOAH phylogeny separates Lund from the evolutionary sequence of Hs and Hss shown in Fig. 1 Tree 2a shows the phylogeny connected to the arrowhead-marked mtDNA introgression that took place from Hss into Hsnn as shown in Fig. 1. The introgression, which gave rise to the Hsnn* branch in 2a, upsets the agreement between the nuDNA and mtDNA trees by joining Hsnn* to the Hss branch in the mtDNA tree, therewith disrupting the initial mtDNA relationship between Hsnn* and SH-Hsnn, an individual representing a population from the Spanish site of Sima de los Huesos, Atapuerca [18,19,20, 62]. The phylogenetic nature of the arising Hss branch is consistent with the strict limitation of Hsnn, the mtDNA sister-group of Hss, to Eurasia [18,19,20,21] and the mtDNA introgression that took place from Hss into Hsnn*, the main branch of Hsnn, ≈ 500,000 YBP, circumstances that are in accordance with OOEH at the same time as they compromise OOAH. The Hss sampling contributing to tree 2a was restricted to the African Mbuti and the Eurasian Lund, two taxa that represent each of the basal lineages among extant humans. This initial representation was extended by the addition of a San and an initial Yoruba to the PPA sampling in tree 2b. The extension joined San and Mbuti on a common branch while the Yoruba joined Lund on the parallel basal Hss branch. In the following PPA step, 2c, the remaining Yorubas were added to the sampling. These Yorubas divided between Lund and the first Yoruba, therewith underlining the separate positions of the Yorubas on the non-African (Lund) branch and the paraphyly of the African Hss populations. The tree in 2d demonstrates the interpretation of Hss relationships and evolution in accordance with OOAH. As apparent this tree, with its root placed in Africa and without connection to its sister-group, Hsn (Hsnn + Hsnd), is refuted both phylogenetically and palaeontologically. The tree is similarly incompatible with the basal nuDNA divergence between Mbuti/San and non-Africans [7, 8]. |
|
|
|
Post by Admin on Jun 10, 2022 17:31:41 GMT
The phylogenetic signification of the difference between the nuDNA and mtDNA trees of Hs The tree in Fig. 1 shows the relationship of Hss and Hsn (Hsnn + Hsnd) consistent with the nuDNA monophyly of Hss and Hsn. In the light of this relationship the divergence between Hss and Hsn ≈ 800,000 YBP becomes anchored in Eurasia in accordance with the principle of Last common ancestor and the strict limitation of Hsn to this continent as maintained earlier [6, 7] and consistent with the recent Ha results [66, 73]. The Hsn branch divides into an Hsnd branch and an Hsnn branch that splits further into SH-Hsnn [18,19,20, 62], and the remaining Hsnn [11, 14, 15], labelled Hsnn*. The arrowheads in the figure mark phylogenetically the mtDNA introgression that took place from Hss into Hsnn* after the divergence between SH-Hsnn and Hsnn*. The mtDNA tree of Hs, Fig. 3, differs fundamentally from the nuDNA phylogeny, Fig. 1, in that Hsnd and SH-Hsnn have become the sole Hsn representatives on a branch (SH-Hsnn + Hsnd) that is sister to the branch that harbours both Hss and Hsnn* as the consequence of the mtDNA introgression that took place from Hss into Hsnn* as marked in Fig. 1. The position of the introgression is consistent with a phylogeny that places the transfer after the Hsnn divergence between SH-Hsnn and Hsnn*. As phylogenetically apparent an introgression in the opposite direction, i.e. from the Hsnn* branch into Hss, would join Hss and Hsnn* on a branch that was sister to SH-Hsnn. Fig. 3 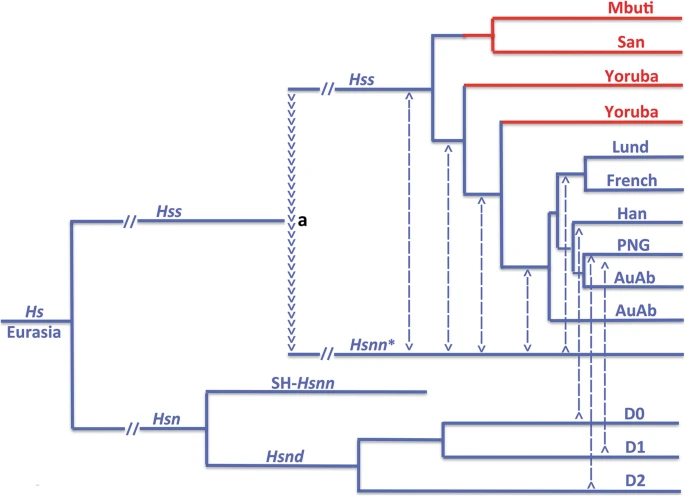 Hs mtDNA relationships. Blue: non-African taxa; red: African taxa. The arrowheads (a) signify the mtDNA introgression from Hss into Hsnn* ≈ 500,000 YBP, after the Hsnn divergence between Hsnn* and SH-Hsnn (see Fig. 1). The Hss phylogeny underlines the evolutionary continuity leading to the non-African populations and the paraphyly of the African populations [7]. The Hsn branch included only one Denisovan in the analysis. Dashed lines indicate general genetic exchanges [11,12,13,14,15] with these from D0, D1 and D2 showing specific input from separate Denisova lineages [78]. The phylogenetic relationship related to SH-Hsnn and the Denisovans is in accordance with independent findings [11, 14, 15, 76, 78,79,80]. AuAb: Australian aborigines; PNG: Papua New Guinean; Han: Chinese; Lund and French: Europeans A Pan/Homo divergence at 8 MYBP [81] places the basal mtDNA divergence in Fig. 3, i.e. that between the branch of (Hss + Hsnn*) and that of (Hsnd + SH-Hsnn) at ≈ 800,000 YBP, the mtDNA introgression from Hss into Hsnn* at ≈ 500,000 YBP and the basal divergence among extant humans at ≈ 250,000 YBP. The mtDNA introgression from Hss into Hsnn* substantiates the presence of the two lineages in the same area ≈ 300,000 years after the divergence between Hss and Hsn ≈ 800,000 YBP. With Hsnn strictly limited to Eurasia the coexistence of Hss and Hsnn confirms the restriction of Hss to this continent [6, 7]. These phylogenetic, geographic and palaeontological circumstances together with the paraphyly of the African populations as demonstrated by the PPA results and the identity of the divergence between Ha and Hs are consistent with an evolutionary scenario that anchors the divergence between Hss and Hsn as well as the mtDNA introgression from Hss into Hsnn* in Eurasia, findings that reverse the direction of Hss evolution compared to that assumed by OOAH. |
|

