|
|
Post by Admin on Jun 10, 2022 20:31:32 GMT
The distinction between the mtDNA and nuDNA trees of Hss and Hsnn was addressed in a molecular study [15] in which the authors related the mtDNA identity of Hsnn to an exodus of an African Hss lineage into Eurasia and the introgression of the mtDNA of this lineage into Hsnn, followed by an extinction of the Hss lineage. That scenario is compromised by the phylogenies shown in Figs. 1 and 3 and the extensive and continuous Eurasian palaeontology of both Hsn and Hss connected to these relationships [e.g. 18–26, 29–34, 38–40, 42–47, 51, 52, 56–58, 60–68]. The age and nature of the Hss fossils at Dali [29, 65], New Cave [30], Jinniushan [33] and Xujiayao [56] included in Fig. 4 are of particular interest in this respect as their ages ≈ 250,000–270,000 years underline the Eurasian existence of Hss at a time that precedes or coincides with the basal divergence among the ancestors of recent Hss. Fig. 4 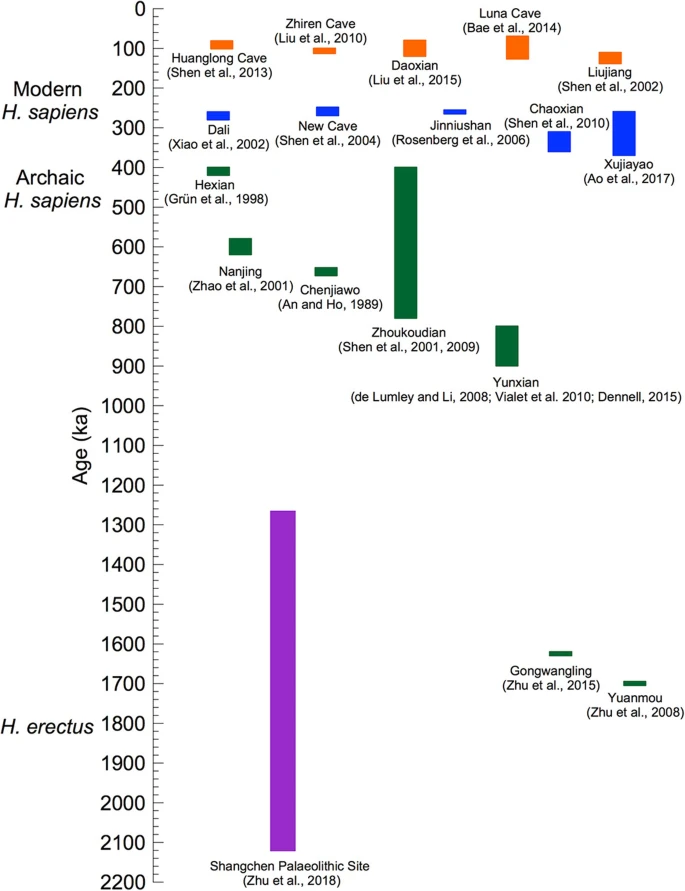 An overview of Chinese palaeontological and archaeological sites related to Hs evolution. The Palaeolithic locality at Shangchen (violet) spans the period from 2,12 to 1,26 MYBP. The Yunxian and Zhoukoudian samplings overlap temporally the divergence into H. antecessor and H. sapiens ≈ 850,000 and Hss and Hsn ≈ 800,000 YBP, making room for that the two lineages arose from a common Eurasian population of H. erectus. The blue section shows more recent Hss positions allowing for admixing with Hsn(Hsnn + Hsnd). The same section covers the interval preceding the divergence between Eurasians and Mbuti/San ≈ 250,000 YBP. The Hualongdong fossils [70], discussed in the text, constitute a part of the blue section. The figure has been adapted and extended from Fig. 10 [56], by permission of the authors In addition to the mtDNA introgression from Hss into Hsnn* ≈ 500,000 YBP Fig. 3 marks general genetic transfers that took place between Hsn (Hsnn + Hsnd) and different Hss populations. The Hsnd input into Papuans was addressed in a study [78] that rested upon OOAH and an African Hss exodus 64,000 YBP, conclusions that differ from the extension of Eurasian palaeontology. The analysis identified two Hsnd introgressions into Papuans, one (D2) at 46,000 YBP the other (D1) at 30,000 YBP. According to the study the D1 branch had split from the Altai Denisovan ≈ 283,000 YBP and the D2 branch ≈ 363,000 YBP. The authors maintained that the D2 estimate was close to that of the divergence between Hsnn and Hsnd, an underestimate of that particular divergence. With respect to the basal Hss divergence among extant humans there is a fundamental phylogenetic discrepancy between Figs. 1, 2 and 3 and the initial studies advocating OOAH [1, 2], which were based on fragmentary mtDNA data. Reanalysis [3] of the data set behind the first study [1] showed that there were thousands of trees that were better by five steps or more than the tree that constituted the basis of the OOAH argument. The terminology used in the study was also inconclusive as the terms “Europa” and “Caucasians” encompassed Europa proper, North Africa and the Middle East (unspecified). Thus, Africa referred only to a part of this continent, compromising the assumption that “Assuming that Africa is the source, there is only one African cluster”. The phylogenetic problems emerged also in the joining of the HeLa lineage (Afro-American) with one Asian, one Australian Aborigine and one European. Similarly, the composite Cambridge mtDNA sequence [82], with its Afro-American HeLa component, grouped among “Europeans”. The follow-up study [2] from the same laboratory was limited to parts of the mtDNA control region. The authors maintained that the analysis had yielded the same pattern as the previous study [1] and that the identification of 14 sequential basal African branches provided the strongest support yet for the placement of the human mtDNA ancestor in Africa. As in the case of the first study a reanalysis of the data [4, 5] identified large numbers of trees that were more parsimonious than the tree that the authors [2] presented and based their OOAH reasoning. |
|
|
|
Post by Admin on Jun 11, 2022 18:27:18 GMT
OOEH and OOAH in the light of palaeontology
The fossil record of Hss is commonly interpreted in compliance with the preconceptions of OOAH in the discussion of Hss evolution. The characterization of the African fossils has a notably wide span, ranging from “The fossil evidence for an African origin for modern humans is robust” [83] to “The hominin fossil record of the African Middle Pleistocene is extremely sparse” [84]. The latter and more inclusive study also detailed that the African fossils were missing adequate provenance in most cases (see Table 1 and Fig. 1 of the study), with the possible exception of the South African Florisbad skull and the age, 260,000 +/− 35,000 years, of the tooth associated with it [85]. Furthermore, Africa contains no fossils of Hsn the mtDNA sister group of Hss, a crucial phylogenetic circumstance as underlined previously [6, 7]. In comparison the limitation of Hsn to Eurasia and the genetic exchanges between Hsn and Hss provide fundamental palaeontological and molecular results that are consistent with the origin and continuous existence of Hss in Eurasia, in accordance with the phylogenies in Figs. 1, 2 and 3.
With respect to the palaeontology and archaeology related to Hss evolution it of particular interest to consider the Eurasian advances in this field during the last 20–25 years. The early stages of the progress were discussed in a study [31] that related the morphological skull mosaic between Hss and H. erectus to a continuous Hs admixing in Asia, including gene flow between eastern and more westerly Asia. As regards the morphological distinction between sapiens and erectus the author [31] recognized them as subspecies, Homo sapiens sapiens and Homo sapiens erectus, in accordance with their overlapping morphology.
The expansion of Eurasian palaeontology has provided insight into the interwoven evolution of H. erectus and Hss in Eastern Eurasia [e.g. 23, 26, 29–31, 33, 40, 41, 45, 50, 51, 56, 61, 63, 64, 67, 69, 86]. One of these studies [56] included a figure, modified here as Fig. 4, which showed the chronostratigraphy among a number of Chinese sites that encompassed H. erectus, archaic and modern Hss and potential Hsnn and Hsnd. The temporal span of the palaeontological findings extended from 1.7 MYBP to 60,000 YBP. The palaeontological and archaeological site at Xujiayao, which was the primary subject of the study [56], covered the period from ≈ 370,000 YBP to ≈ 250,000 YBP, i.e. a substantial part of Hss evolution prior to the Hss divergence between Mbuti/San and the branch of other extant humans as shown in Figs. 1, 2 and 3. The Xujiayao site belongs to the particular temporal span (blue) that extends from ≈ 380,000 YBP to ≈ 220,000 YBP in Fig. 4, which the authors [56] related to the late coexistence and genetic admixture of H. erectus and early Hss plus an Hsn input from potentially both Hsnn and Hsnd. It should be noted that the latter part of this timespan coincides with the genetic exchanges that have been recorded between Hsnn and Hss before the basal divergence among recent humans [12, 13, 15,16,17].
The early part of the time range of Xujiayao coincides temporally with the Chaoxian site [40] while the more recent range of the site overlaps with the age of the sites at Dali [29, 65], New Cave [30], Jinniushan [33] and Hualongdong [70], which also provide palaeontological evidence for an extended Hsn/Hss interface in Eurasia preceding the basal Hss divergence between the ancestors of Mbuti/San and remaining Hss populations shown in Figs. 1, 2 and 3. Regarding the volume of the samples from the Xujiayao site it may be noted that they have, in addition to 20 fossil human specimens, yielded more than 30,000 lithic artefacts and about 5000 mammalian fossils that substantiate the nature and age and of the human specimens.
With respect to the age, 1,7 MY, of the Yuanmou finds [36] and that of 1,63 MY of the Gongwangling samples [54] in Fig. 4 it is of crucial interest that their age became exceeded by a wide margin by the artefact sequence from Shangchen, near the Gongwangling site [68]. The 17 continuous artefact layers studied covered a timespan from 2,12 MYBP to 1,26 MYBP. The authors concluded that the findings were in accordance with hominins leaving Africa considerable earlier than indicated by the age, 1,77–1,85 MY, of the Homo erectus fossils from the Georgian Dmanisi site [87, 88]. The easterly range of H. erectus is represented by the younger Indonesian distribution of the taxon [89,90,91].
Archaeological sites east of the Mediterranean Sea have revealed a human presence as early as 400,000 YBP [25, 38]. The fossils were not assigned specifically to either Hss or Hsn, however. In comparison, the human fossils from Qafzeh and Skhul with an age of 90,000–100,000 years were identified as distinct Hss [23, 26]. These fossils together with later described Hss fossils from the same region, with an age of 180,000–200,000 years [22] fall into the western part of the Eurasian Hss distribution that corresponds to the span of modern H. sapiens in Fig. 4, i.e. Hss after the divergence between Mbuti/San and the remaining Eurasian populations.
The African palaeontology related to Hs origin and evolution is distinctly poorer than that of Eurasia. The African Homo picture outside Hss became extended significantly, however, with the description of the fossils of H. naledi, which comprise by far the largest assemblage of Homo fossils in Africa. The fossils come from two sites in the South African Rising Star cave system, the Dinaledi Chamber and the Lesedi Chamber [84, 92]. Neither site contains other Homo fossils. Dating of the Dinaledi fossils constrains their age to 236,000–335,000 YBP. Another estimate based on the two least-altered naledi teeth found yielded a maximum average age of 253 + 82/− 70 thousand years and a minimum average age of 200 + 70/− 61 thousand years [93].
|
|
|
|
Post by Admin on Jun 11, 2022 21:29:21 GMT
The extensiveness of the H. naledi fossils is notable compared to the scarcity of Hss fossils in a region that according to proponents of OOAH may have constituted the cradle of Hss. Another circumstance relates to the question how Hss and H. naledi, which might have shared similar ecological niches, could thrive contemporaneously in the same area. Taken together the findings allow the proposal that naledi evolved in southern Africa without competition from Hss and that it was a later intrusion of Hss from the north that led to the demise of H. naledi. In this case the Florisbad fossil would constitute a representative of early Hss intruders (possibly Mbuti/San) coming from the north.
The redating of the fossils at the Jebel Irhoud site, Morocco [94, 95], which increased their age by ≈ 100,000 years compared to an earlier study of the same site [96], renders the fossils of interest for the discussion of Hss evolution in the light of the deepest divergence among recent humans, ≈ 250,000 YBP, and the reversal of the direction of evolution of the Hs tree as discussed in connection with Figs. 1, 2 and 3. The age of the tooth examined was 286+/− 32 thousand years, somewhat younger than the age, 315+/− 34 thousand years, of the artifacts connected to layer 7 [94, 95]. The finds have particular implications in the light of the Chinese finds of Dali, New Cave, Jinniushan, Xujiayao and Hualongdong, blue in Fig. 4, which fall in the same part of the temporal interval between the mtDNA introgression from Hss into Hsnn* ≈ 500,000 YBP and the basal divergence among recent Hss ≈ 250,000 YBP. As the revised age of the Jebel Irhoud fossils exceeds by some margin the age of the basal divergence among extant humans it appears that the Jebel Irhoud population became extinct without contributing to the genetic constitution of extant Africans.
A recent study [97] of the Greek palaeontological finds Apidima 1 and Apidima 2 characterized the Apidima 1 fossil with an allocated age of ≈ 210,000 years as constituting the earliest known presence of Homo sapiens (Hss here) in Eurasia. This conclusion is implausible considering both the calculation of the age of the Apidima 1 fossil [71, 98] and the large series of much older Eurasian palaeontological finds related to Hss evolution that had escaped the attention of the authors [97]. A similar disregard of Eurasian palaeontology appeared in a preceding study [99] that detailed a scenario that rested upon an African origin and divergence between Neanderthals and Denisovans ≈ 480,000 YBP and their exodus into Eurasia ≈ 400,000 followed by an Out of Africa dispersal of modern humans ≈ 75,000–125,000 YBP, circumstances that are incompatible with the extensive Eurasian palaeontology of both Hsn and Hss and molecular findings, including the molecular exchanges between Hss and Hsn (Hsnn + Hsnd). Similarly, a more comprehensive study [100] that appeared in the same journal did not allow a treatment of the Eurasian palaeontology related to Hss origin and dispersal as the discussion rested upon OOAH and a temporal demarcation at 125,000 YBP for the oldest Eurasian fossils discussed in the context of Hss evolution.
|
|
|
|
Post by Admin on Jun 12, 2022 19:30:20 GMT
The OOEH synthesis The present reorientation of the Hs tree in conjunction with the palaeontology of H. erectus makes the period ≥900,000 to 500,000 YBP highly significant for the discussion of Hs evolution as it covers both the advent and sequel of the divergences that encompass H. erectus, H. antecessor, Hss and Hsn (Hsnn + Hsnd) shown in Fig. 1. The Asian phase of H. erectus evolution ≥900,000 YBP is exemplified in Fig. 4 by the Chinese site at Yunxian [35, 41, 53, 56] and the age, 936,000 years, of the best-preserved scull (Yunxian II) excavated at this site [35, 41]. With regard to the Yunxian II specimen one of these studies [41] concluded that the facial features of the fossil displayed a pattern close to modern humans and that the assignment of the scull to H. erectus extended the variability connected to this species. In this light and in recognition of the fossil record of H. erectus the findings are consistent with the origin of a branch that arose within a diversifying Asian H. erectus population which diverged into the westward going population of H. antecessor, which came to reside for a short period in SW Europa, and another population, H. sapiens, that split later into Hss and Hsn. According to this understanding Hsn diverged further into Hsnd and Hsnn with Hsnd diversifying essentially in eastern Asia and the Sahul and Hsnn inhabiting a large Eurasian area within which it diversified into a westerly branch, SH-Hsnn, and a more widely spread population, Hsnn*, characterized by the mtDNA introgression from Hss ≈ 500,000 YBP, a circumstance that manifests the lasting and contemporary Eurasian presence of Hss and Hsnn. It should be noted that the Hss part of the mtDNA tree in Fig. 3 suggests that the Eurasian Hss population went through a severe bottleneck prior to the diversification shown in the figure. In comparison the African population structure suggests that the populations arising from the different Hss exoduses into Africa remained relatively unaffected in this respect. Hss evolution in relation to climatic changes The estimated ages of the divergences related to Hss and Hsn show considerable variation among authors depending on the calibration points chosen. In the present study we have applied as a calibration point the Homo/Pan divergence set at 8 MYBP, in accordance with the mammalian calibration point A/C-60 [81]. As mentioned above the 8 MY age of the Homo/Pan divergence places the basal Hs divergence between Hss and Hsn at ≈ 800,000 YBP and the mtDNA introgression from Hss into Hsnn* at ≈ 500,000 YBP. Within the Hss lineage itself the basal divergence, that between Mbuti/San and other recent lineages, becomes placed at ≈ 250,000 YBP, the time of the two Yoruba exoduses into Africa at ≈ 225,000 and ≈ 180,000 YBP respectively and the age of the basal divergence among recent non-Africans at ≈ 125,000 YBP. The ages of three of these estimates, ≈ 250,000, ≈ 225,000 and ≈ 125,000 YBP, coincide with warm global temperatures that were preceded by low maxima as recorded in analyses of Antarctic ice core records [101,102,103]. The picture is consistent with a Eurasian Hss population that passed three cold-related bottlenecks, each of which was followed by population expansion and concomitant dispersal. Although the coincidence between the timings of the molecular estimates and climatic changes might not be absolute they underline the scenario of oscillating climatic conditions that are likely to have affected vegetation and population structures among both humans and their prey in connection with changes in sea levels and routes of dispersal. The molecular variation within Africa in both mtDNA [104] and nuDNA [8] is greater than that among non-African populations. Among population geneticists this variation has been commonly interpreted as supporting OOAH without considering the phylogenetic nature of the relationships connected to the different Hs divergences leading to recent humans. The identity of these relationships as maintained here and the paraphyly of the African populations suggests a different explanation, namely that the African variation is the effect of consecutive Hss dispersals from Eurasia into Africa and climate-related Eurasian Hss bottlenecks that reduced the non-African variation, the most distinct being that ending ≈ 125,000 YBP. A scenario of this kind concurs with climate history and the glaciation cycles discussed above [101,102,103]. It has been noted similarly in several studies [17, 105,106,107] that the amount of Hsnn DNA in African genomes is less than that of non-Africans, a finding that is consistent with the African absence of both Neanderthals and Denisovans and an extended contact by Neanderthals and Denisovans with the non-African populations. Figure 5 shows potential routes related to the dispersal of Hss in accordance with the PPA findings and the relationships behind Figs. 1, 2 and 3. The depicted routes allow ample room for the genetic exchanges between Hss and Hsnn and Hss and Hsnd in Eurasia and more south-easterly regions that have been detailed in a series of nuDNA studies [e.g. 10, 12, 13, 15–17, 80, 81, 108–111]. 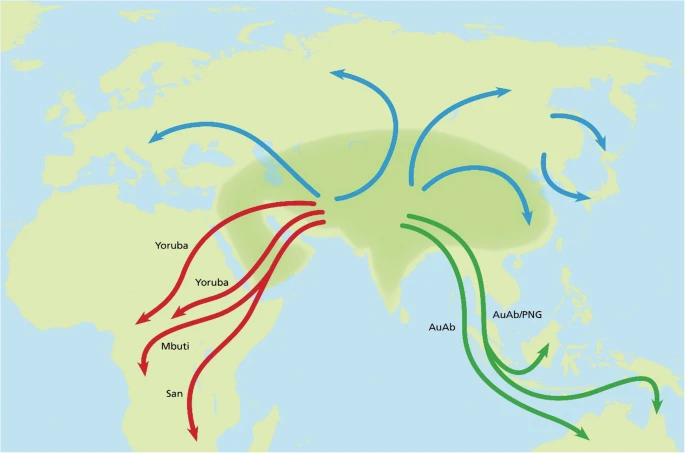 Fig. 5 A simplified view of Hss dispersal. The shaded area signifies a geographically undefined Asian (Eurasian) area from which Hss dispersed. Mbuti/San mark the earliest Hss exodus into Africa followed by later Yoruba exoduses. The green tracks represent routes that signify potential routes into Southeast Asia and Oceania |
|
|
|
Post by Admin on Jun 12, 2022 21:56:25 GMT
Conclusions
The reversal of the direction of evolution in the tree of Homo sapiens sapiens is consistent with the molecular identification of the sister group relationship between H. sapiens and H. antecessor and a Eurasian separation into H. antecessor and H. sapiens (Hss + Hsn) ≈ 850.000 YBP [66, 73] within a diversifying population of H. erectus. The hitherto earliest recorded phase of hominin presence in Eurasia, that at Shangchen, China, has been dated to ≈ 2.1 MYBP [68]. The finds constitute a temporal extension of the palaeontological findings of H. erectus in Dmanisi, Georgia, ≥ 1.8 MYBP [87, 88] and Yuanmou, China, 1.7 MYBP [36] and a series of younger finds that underline the Eurasian existence of H. erectus preceding the divergence between H. antecessor and H. sapiens.
We postulate that following the separation between H. sapiens and H. antecessor ≈ 850.000, Hs diverged further into Hss and Hsn ≈ 800.000 YBP. The Eurasian continuity of these two lineages is underlined by the molecular diversity of Hsnn and Hsnd and the mtDNA introgression that took place from Hss into Hsnn* ≈ 500,000 YBP. In comparison the Hss branch remains undivided until the basal divergence among the ancestors of extant Hss as represented in Fig. 1 by Lund and the African populations of Mbuti/San, therewith bringing to an end the evolutionary journey that began by Homo erectus leaving Africa ≥2 million years earlier.
|
|

