Post by Admin on Sept 7, 2023 19:23:45 GMT
Fig. 3: Strontium data of pedigree A.
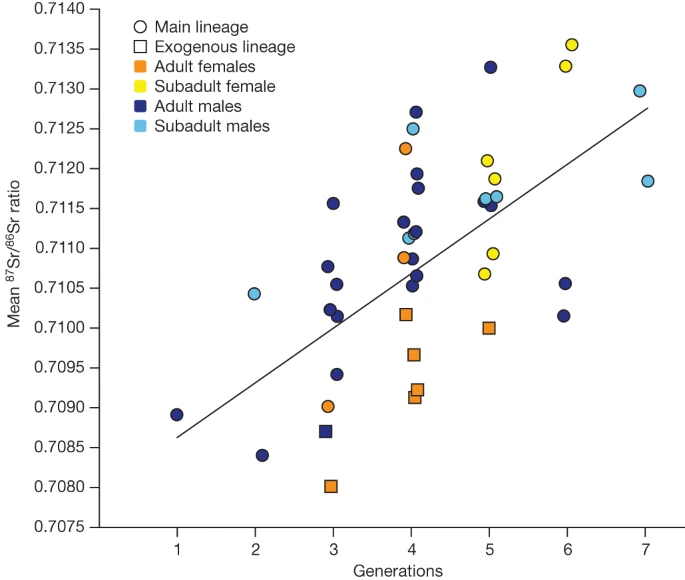
Mean 87Sr/86Sr ratio per age and sex cohort across generations. A significant difference between sex was observed per generation (two-sided analysis of variance, P = 0.01474; Supplementary Note 11).
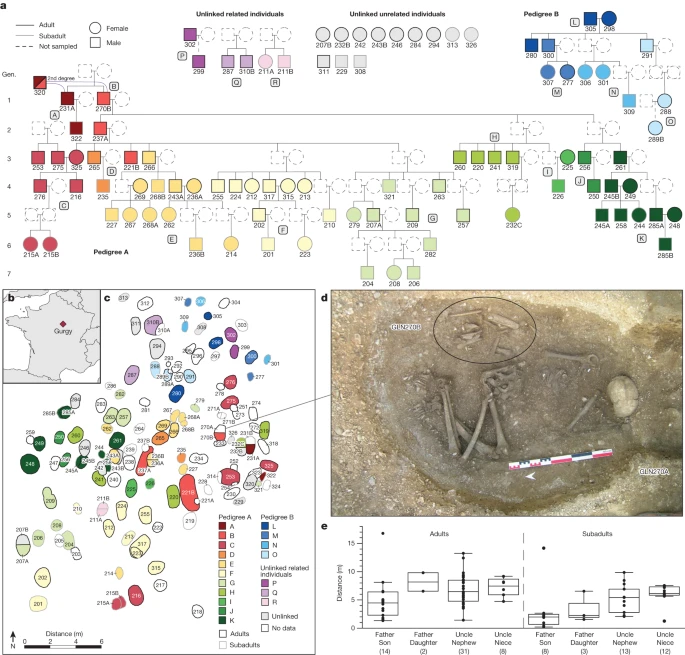
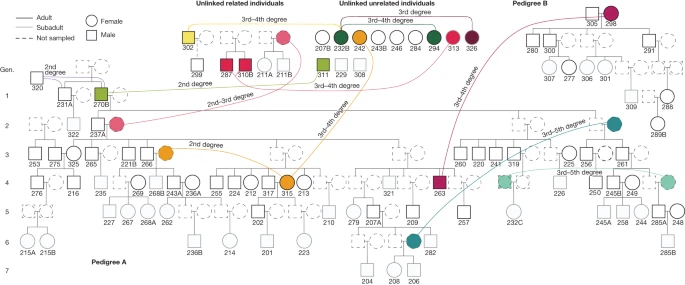
The presumed patrilocal residential pattern in the Gurgy group also explains the mitochondrial diversity despite the deficit in female adults (35 different mitochondrial haplogroups are carried by 99 individuals; Supplementary Note 6, Supplementary Table 5 and Extended Data Fig. 3). Indeed, no mitochondrial haplogroup was transmitted further than one daughter/son generation, and the incoming mothers of each generation contributed new mtDNA lineages, except for the female descendant of the main lineage GLN325 who transmitted hers one generation further. By examining the affinities between all of the exogenous female adults, we demonstrate that they are not closely related, apart from two pairs of third- or fourth-degree relatives (Supplementary Note 5 and Extended Data Fig. 4). Moreover, this female diversity within the Gurgy group might also explain the overall phenotypic variation observed at the site (Supplementary Note 9 and Supplementary Tables 15 and 16). Taken together, these results suggest that the Gurgy community maintained a fairly clear pattern of female exogamy that may have been driven by a range of group features (for example, population size, resource access, network position) or identities (for example, linguistic or cultural affinities). The absence of long runs of homozygosity (ROH), typical for close-kin consanguinity, confirms the avoidance of reproduction between closely related individuals, except for a single individual (GLN282), with an amount of ROH consistent with a union between second- or third-degree relatives (Supplementary Note 4, Supplementary Table 11 and Extended Data Fig. 9). IBD sharing also revealed that groups of individuals in pedigree A are more related than expected from connections within the reconstructed pedigree alone (Fig. 2 and Supplementary Note 3). This can only be explained if there were additional relatives through maternal lines (which were not sampled). In our case, some mothers probably came from the same external group, only several generations apart, which would link different branches of the pedigree. For example, siblings GLN243A and GLN268B share the same mitochondrial haplotype H1 as GLN315 (Supplementary Table 5) and IBD-sharing typical of third-degree relationships, whereas GLN315 is an exogenous female individual. We interpret these as a second-degree relationship between GLN315 and the missing mother of GLN243A and 268B (Fig. 2). These additional connections through the female lines indicate a network of relationships with other groups, including occasional unions with (distantly) related women from the same source group. This pattern suggests preferential links or dependencies between some groups, albeit within a network of groups sufficiently large or diverse to sustain diversity of background relatedness and to avoid close-kin consanguinity.
Further insights about the social organization of the group can be gleaned from the notable lack of half-siblings in the entire sample, in contrast to recent findings from a later Neolithic long cairn in England8 (Supplementary Notes 2 and 3). This indicates that polygamous reproductive unions were uncommon or perhaps socially proscribed, or that the burial of offspring from such unions was carried out elsewhere. Likewise, it also suggests that serial monogamy, including levirate and sororate unions in which a woman repartners with her deceased husband’s brother or a man repartners with his wife’s sister, was rare. We find this observation surprising given potential imbalances in the female/male sex-ratio, for example, an elevated risk of death from complications during childbirth (for female individual), potential conflicts or diseases in prehistoric societies. The pedigrees show no evidence in support of these assumptions. Indeed, if exogamous reproductive unions were routinely contracted with numerous groups for purposes of, for example, alliance or trade, then between-group networks of cooperation, rather than conflict, are implied.
Including unsampled, inferred adults, we observed two cases of up to six offspring from the same couple (Extended Data Fig. 6). Notably, all six full siblings had reached reproductive age, with several of those having four and five adult offspring on their own. Moreover, the majority of adult offspring being male individuals points towards additional unsampled female siblings (to statistically account for an equivalent number of females born19), as well as possibly a significant number of deceased infants expected at that period of time. These large family sizes suggest a high fertility rate and generally stable conditions of health and nutrition in this Neolithic community5—a fact that is also supported by stable isotope data23. Indeed, one could speculate that different elements, such as a potential emphasis on sublineage reproductive and/or productive units, spatial co-residence of numerous reproductive units and divisions of labour that may facilitate efficient reproduction, plausibly provide conditions for cooperative breeding that can generate high rates of population growth24. We estimated the effective population size of the communities contributing to the diversity observed at Gurgy to have been around 1,835 individuals (95% CI = 1,631–2,077)25 (Supplementary Note 4 and Supplementary Table 26). The distribution of ROH in the group (Supplementary Table 11 and Extended Data Fig. 9) suggests that most pairs of parents were related to each other through co-ancestors within the preceding 5–30 generations (Supplementary Note 4).
Mean 87Sr/86Sr ratio per age and sex cohort across generations. A significant difference between sex was observed per generation (two-sided analysis of variance, P = 0.01474; Supplementary Note 11).
The presumed patrilocal residential pattern in the Gurgy group also explains the mitochondrial diversity despite the deficit in female adults (35 different mitochondrial haplogroups are carried by 99 individuals; Supplementary Note 6, Supplementary Table 5 and Extended Data Fig. 3). Indeed, no mitochondrial haplogroup was transmitted further than one daughter/son generation, and the incoming mothers of each generation contributed new mtDNA lineages, except for the female descendant of the main lineage GLN325 who transmitted hers one generation further. By examining the affinities between all of the exogenous female adults, we demonstrate that they are not closely related, apart from two pairs of third- or fourth-degree relatives (Supplementary Note 5 and Extended Data Fig. 4). Moreover, this female diversity within the Gurgy group might also explain the overall phenotypic variation observed at the site (Supplementary Note 9 and Supplementary Tables 15 and 16). Taken together, these results suggest that the Gurgy community maintained a fairly clear pattern of female exogamy that may have been driven by a range of group features (for example, population size, resource access, network position) or identities (for example, linguistic or cultural affinities). The absence of long runs of homozygosity (ROH), typical for close-kin consanguinity, confirms the avoidance of reproduction between closely related individuals, except for a single individual (GLN282), with an amount of ROH consistent with a union between second- or third-degree relatives (Supplementary Note 4, Supplementary Table 11 and Extended Data Fig. 9). IBD sharing also revealed that groups of individuals in pedigree A are more related than expected from connections within the reconstructed pedigree alone (Fig. 2 and Supplementary Note 3). This can only be explained if there were additional relatives through maternal lines (which were not sampled). In our case, some mothers probably came from the same external group, only several generations apart, which would link different branches of the pedigree. For example, siblings GLN243A and GLN268B share the same mitochondrial haplotype H1 as GLN315 (Supplementary Table 5) and IBD-sharing typical of third-degree relationships, whereas GLN315 is an exogenous female individual. We interpret these as a second-degree relationship between GLN315 and the missing mother of GLN243A and 268B (Fig. 2). These additional connections through the female lines indicate a network of relationships with other groups, including occasional unions with (distantly) related women from the same source group. This pattern suggests preferential links or dependencies between some groups, albeit within a network of groups sufficiently large or diverse to sustain diversity of background relatedness and to avoid close-kin consanguinity.
Further insights about the social organization of the group can be gleaned from the notable lack of half-siblings in the entire sample, in contrast to recent findings from a later Neolithic long cairn in England8 (Supplementary Notes 2 and 3). This indicates that polygamous reproductive unions were uncommon or perhaps socially proscribed, or that the burial of offspring from such unions was carried out elsewhere. Likewise, it also suggests that serial monogamy, including levirate and sororate unions in which a woman repartners with her deceased husband’s brother or a man repartners with his wife’s sister, was rare. We find this observation surprising given potential imbalances in the female/male sex-ratio, for example, an elevated risk of death from complications during childbirth (for female individual), potential conflicts or diseases in prehistoric societies. The pedigrees show no evidence in support of these assumptions. Indeed, if exogamous reproductive unions were routinely contracted with numerous groups for purposes of, for example, alliance or trade, then between-group networks of cooperation, rather than conflict, are implied.
Including unsampled, inferred adults, we observed two cases of up to six offspring from the same couple (Extended Data Fig. 6). Notably, all six full siblings had reached reproductive age, with several of those having four and five adult offspring on their own. Moreover, the majority of adult offspring being male individuals points towards additional unsampled female siblings (to statistically account for an equivalent number of females born19), as well as possibly a significant number of deceased infants expected at that period of time. These large family sizes suggest a high fertility rate and generally stable conditions of health and nutrition in this Neolithic community5—a fact that is also supported by stable isotope data23. Indeed, one could speculate that different elements, such as a potential emphasis on sublineage reproductive and/or productive units, spatial co-residence of numerous reproductive units and divisions of labour that may facilitate efficient reproduction, plausibly provide conditions for cooperative breeding that can generate high rates of population growth24. We estimated the effective population size of the communities contributing to the diversity observed at Gurgy to have been around 1,835 individuals (95% CI = 1,631–2,077)25 (Supplementary Note 4 and Supplementary Table 26). The distribution of ROH in the group (Supplementary Table 11 and Extended Data Fig. 9) suggests that most pairs of parents were related to each other through co-ancestors within the preceding 5–30 generations (Supplementary Note 4).