Post by Admin on Dec 27, 2023 21:30:13 GMT
Social relationships and residence
Examining both pedigrees, we found that generations are linked almost exclusively through the male line, that is, all descendants but one are connected to the family tree through their father’s line. The Y-chromosome haplogroup G2a2b2a1a2 (terminal SNP Z38302) is carried by 51 out of 57 male individuals and is the main male lineage of the group (Supplementary Table 7 and Extended Data Fig. 2). Pedigree A yields the sole exception, where a new lineage (C in Fig. 1a) is linked through a lineage woman (GLN325). Her reproductive partner and his brother, her sons and two other unrelated male individuals carry haplogroup H2m, the only other Y-chromosome lineage observed in the dataset (Fig. 1a and Extended Data Fig. 2). The main genealogical lineage of pedigree A is also visible in the funerary features, as one son, GLN237A, and one grandson, GLN221B, of the main ancestor GLN270B are buried in the largest pits of the necropolis (Fig. 1a, Supplementary Note 1 and Extended Data Fig. 6). GLN270B, the main ancestor of 52 individuals in pedigree A, represents the only secondary burial at the site, consisting of long bones that were buried together with the female individual GLN270A, for whom no genomic data could be obtained (Fig. 1d). The position at the apex of the pedigree suggests that his remains were transferred and buried during the early occupation of the site, representing, together with his brother GLN231A, the main male posthumous ancestors of pedigree A. The association between GLN270B and female individual GLN270A, and not with his brother, for example, suggests that this female individual was important—perhaps she was his partner, or someone genetically related representing the lineage.
When we investigated the spatial organization of the graves by measuring the physical distance between individuals, we observed significantly closer spatial proximity between each father–son pair than any other related pair (Fig. 1e, Supplementary Note 12 and Supplementary Table 20). Patterns of spatial organization beyond the specific father–son connections (Supplementary Note 12) seem to follow clusters of genetically closely related individuals (Fig. 1a,c). Indeed, after an initial phase of early burials that were grouped together in the eastern part of the funerary area, the siblings from the fourth generation were all buried near each other (Fig. 1a,c). For example, the four siblings of individual GLN317 were buried west of him, whereas the mother of his sons was buried east of him, and GLN223 and his son’s daughter was buried on top of him. The other son of GLN317, GLN202, probably died later as he was buried in another part of the necropolis, together with other branches of pedigree A, and was possibly the most recently deceased of this family line. We co-analysed the spatial distribution of the few grave goods and adornments attributed to the dead, such as ornaments or ochre, and potential transmission along the paternal lineage using a Geographic Information System (Supplementary Note 12 and Extended Data Figs. 3 and 6). However, perhaps due to the general paucity of grave goods at the site (Supplementary Table 21), no association was detected. We also did not find any correlations between specific body positions (crouched/flexed or elongated), sidedness and orientation, type of grave and genetic lineages, genetic ‘nuclear’ families (that is, mother–father–child trios), and/or unlinked/unrelated individuals (Supplementary Note 12 and Extended Data Fig. 3). However, the spatial layout with minimal or no overlap between burials suggests that graves were visible or marked on the surface18, and the pattern of expansion indicates that people knew who was buried where, and may have acknowledged lines of descent accordingly. Together, we observed a general trend at the biological and archaeological level, in which individuals are linked through the male lineage, potentially indicative of local understandings of genealogy or descent.
The pedigree structure of the burial community reveals further insights into the residential organization of the living. Apart from two individuals (GLN325 (see above) and GLN288), no adult mothers, present (n = 7) or absent, have parents/ancestors buried at the site. This suggests an exogenous origin of these females (Extended Data Figs. 2 and 7). Moreover, only 6 out of the 20 female adults buried at the site are descendants of the main pedigree lines A and B. Another seven female adults buried at Gurgy had very few biological relationships with other individuals, and mainly not of the main pedigrees, as shown by IBD analyses (Fig. 2). One plausible hypothesis is that they were companions of male individuals from the main pedigrees: no joint children were buried on site, nor could they be linked through other individuals to the pedigrees. Indeed, 17 adult male individuals have no children buried at the site, of which 13 are linked through their parents to either main pedigree. This general pattern points towards female exogamy and a virilocal residential system in which females in-migrated from their birthplace to their male reproductive partner’s residence. Consistent with this pattern, additional links observed between the isolated female adults and the pedigrees could be due to (1) distantly related female individuals stemming from the same community; or (2) women who left the Gurgy community in previous generations with female descendants who subsequently returned to Gurgy. The latter scenario is indicative of reciprocal exchange typical in moiety systems3. Perhaps as a consequence, the sex ratio of adult offspring buried on site is unbalanced at 4.5:1 (confidence interval (CI) = 64.5–93%) in favour of male individuals (n = 27 versus n = 6, for male and female, respectively). By contrast, a sex ratio of 1.06:1 (CI = 34.4–68.1%) among subadult offspring (n = 19 and n = 18, for male and female, respectively) matches the natural expected ratio 1.05:1 at birth19, ruling out sex-biased cultural practices affecting the subadult population (Extended Data Fig. 2). The vast majority of the subadults are younger than 15 years old (n = 34), with most of these individuals being younger than 8 years old (n = 27), and in equal proportions for both sexes. The differences between younger and older-age sex ratios suggest that older daughters, from around the age of 15, left to join new groups, again consistent with a female exogamic residential system. For four out of the six adult lineage daughters (GLN212, 213, 277 and 289B) who remained at Gurgy, no offspring could be identified at the site even though they had reached reproductive age. Female exogamy may not have been practiced strictly or, alternatively, these lineage daughters could be reproductive partners of unlinked adult males (with no offspring linking them to the pedigrees)—a scenario that further complicates the assumption of strict patrilocality and female exogamy. Alternative reasons for their stay in the community remain unclear.
In this context, we observed that women of genetically exogenous provenance tend to be spatially integrated into their reproductive partner’s burial area, suggesting social integration into the host group (Fig. 1a). However, considering the 42 reproductive unions observed across all pedigrees that evidently had offspring buried at Gurgy, we noticed a shortage of mothers, with only 9 versus 20 fathers buried there. This imbalance is also observed in the total number of adult burials (38 male versus 20 female), and suggests that male adults were twice as likely to be buried than female adults. We therefore observed a potential sex bias in burials independent of female exogamy. This could be explained by different funerary practices being reserved for these mothers, or by other social factors mitigating against a co-burial with their reproductive partner’s group.
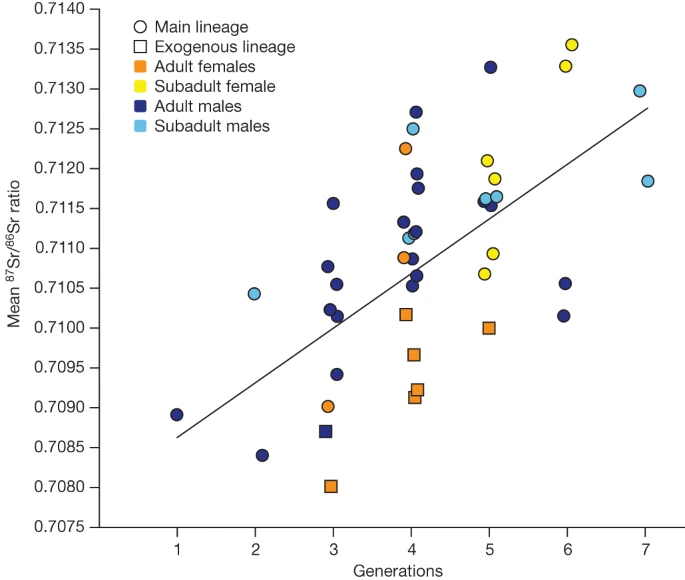
To gain independent information on individual mobility, we performed strontium isotope analyses (87Sr/86Sr) using laser ablation20 on 57 individuals (Fig. 3, Supplementary Note 11, Supplementary Table 23 and Extended Data Figs. 3 and 8). Unrelated female adults and some of those with no parents at the site show lower 87Sr/86Sr ratio values compared with male individuals from the same generation (Fig. 3 and Extended Data Fig. 3). Although the geological reference map does not enable us to infer a specific geographical origin, this finding provides further evidence that these female individuals grew up in different places before joining the Gurgy community (Supplementary Note 11 and Extended Data Fig. 8). Published stable isotope data (carbon, nitrogen, sulphur) measured on bones highlight a significant, sex-biased dietary division in adults21. On average, male individuals yielded higher δ13C and δ15N, and lower δ34S values, than female individuals, which could reflect a separation by sex, but could also signal female mobility (Supplementary Table 24). Genetic sex determination of subadults enabled us to confirm this difference also in childhood (P = 0.01019), which could be explained by a sex-related differential treatment at certain ages, determined by social rules (Supplementary Note 14 and Extended Data Fig. 5). Notably, the funerary practices in Gurgy show a shift at around 7–8 years of age, when the children are buried with different types of grave goods compared with younger ages, and another shift at around 15–16 years of age when they are associated with the same grave goods as adults, which could reflect local age stages or other social thresholds. This pattern has previously been observed at other Neolithic sites in the northern half of France22 (Supplementary Table 1).
Examining both pedigrees, we found that generations are linked almost exclusively through the male line, that is, all descendants but one are connected to the family tree through their father’s line. The Y-chromosome haplogroup G2a2b2a1a2 (terminal SNP Z38302) is carried by 51 out of 57 male individuals and is the main male lineage of the group (Supplementary Table 7 and Extended Data Fig. 2). Pedigree A yields the sole exception, where a new lineage (C in Fig. 1a) is linked through a lineage woman (GLN325). Her reproductive partner and his brother, her sons and two other unrelated male individuals carry haplogroup H2m, the only other Y-chromosome lineage observed in the dataset (Fig. 1a and Extended Data Fig. 2). The main genealogical lineage of pedigree A is also visible in the funerary features, as one son, GLN237A, and one grandson, GLN221B, of the main ancestor GLN270B are buried in the largest pits of the necropolis (Fig. 1a, Supplementary Note 1 and Extended Data Fig. 6). GLN270B, the main ancestor of 52 individuals in pedigree A, represents the only secondary burial at the site, consisting of long bones that were buried together with the female individual GLN270A, for whom no genomic data could be obtained (Fig. 1d). The position at the apex of the pedigree suggests that his remains were transferred and buried during the early occupation of the site, representing, together with his brother GLN231A, the main male posthumous ancestors of pedigree A. The association between GLN270B and female individual GLN270A, and not with his brother, for example, suggests that this female individual was important—perhaps she was his partner, or someone genetically related representing the lineage.
When we investigated the spatial organization of the graves by measuring the physical distance between individuals, we observed significantly closer spatial proximity between each father–son pair than any other related pair (Fig. 1e, Supplementary Note 12 and Supplementary Table 20). Patterns of spatial organization beyond the specific father–son connections (Supplementary Note 12) seem to follow clusters of genetically closely related individuals (Fig. 1a,c). Indeed, after an initial phase of early burials that were grouped together in the eastern part of the funerary area, the siblings from the fourth generation were all buried near each other (Fig. 1a,c). For example, the four siblings of individual GLN317 were buried west of him, whereas the mother of his sons was buried east of him, and GLN223 and his son’s daughter was buried on top of him. The other son of GLN317, GLN202, probably died later as he was buried in another part of the necropolis, together with other branches of pedigree A, and was possibly the most recently deceased of this family line. We co-analysed the spatial distribution of the few grave goods and adornments attributed to the dead, such as ornaments or ochre, and potential transmission along the paternal lineage using a Geographic Information System (Supplementary Note 12 and Extended Data Figs. 3 and 6). However, perhaps due to the general paucity of grave goods at the site (Supplementary Table 21), no association was detected. We also did not find any correlations between specific body positions (crouched/flexed or elongated), sidedness and orientation, type of grave and genetic lineages, genetic ‘nuclear’ families (that is, mother–father–child trios), and/or unlinked/unrelated individuals (Supplementary Note 12 and Extended Data Fig. 3). However, the spatial layout with minimal or no overlap between burials suggests that graves were visible or marked on the surface18, and the pattern of expansion indicates that people knew who was buried where, and may have acknowledged lines of descent accordingly. Together, we observed a general trend at the biological and archaeological level, in which individuals are linked through the male lineage, potentially indicative of local understandings of genealogy or descent.
The pedigree structure of the burial community reveals further insights into the residential organization of the living. Apart from two individuals (GLN325 (see above) and GLN288), no adult mothers, present (n = 7) or absent, have parents/ancestors buried at the site. This suggests an exogenous origin of these females (Extended Data Figs. 2 and 7). Moreover, only 6 out of the 20 female adults buried at the site are descendants of the main pedigree lines A and B. Another seven female adults buried at Gurgy had very few biological relationships with other individuals, and mainly not of the main pedigrees, as shown by IBD analyses (Fig. 2). One plausible hypothesis is that they were companions of male individuals from the main pedigrees: no joint children were buried on site, nor could they be linked through other individuals to the pedigrees. Indeed, 17 adult male individuals have no children buried at the site, of which 13 are linked through their parents to either main pedigree. This general pattern points towards female exogamy and a virilocal residential system in which females in-migrated from their birthplace to their male reproductive partner’s residence. Consistent with this pattern, additional links observed between the isolated female adults and the pedigrees could be due to (1) distantly related female individuals stemming from the same community; or (2) women who left the Gurgy community in previous generations with female descendants who subsequently returned to Gurgy. The latter scenario is indicative of reciprocal exchange typical in moiety systems3. Perhaps as a consequence, the sex ratio of adult offspring buried on site is unbalanced at 4.5:1 (confidence interval (CI) = 64.5–93%) in favour of male individuals (n = 27 versus n = 6, for male and female, respectively). By contrast, a sex ratio of 1.06:1 (CI = 34.4–68.1%) among subadult offspring (n = 19 and n = 18, for male and female, respectively) matches the natural expected ratio 1.05:1 at birth19, ruling out sex-biased cultural practices affecting the subadult population (Extended Data Fig. 2). The vast majority of the subadults are younger than 15 years old (n = 34), with most of these individuals being younger than 8 years old (n = 27), and in equal proportions for both sexes. The differences between younger and older-age sex ratios suggest that older daughters, from around the age of 15, left to join new groups, again consistent with a female exogamic residential system. For four out of the six adult lineage daughters (GLN212, 213, 277 and 289B) who remained at Gurgy, no offspring could be identified at the site even though they had reached reproductive age. Female exogamy may not have been practiced strictly or, alternatively, these lineage daughters could be reproductive partners of unlinked adult males (with no offspring linking them to the pedigrees)—a scenario that further complicates the assumption of strict patrilocality and female exogamy. Alternative reasons for their stay in the community remain unclear.
In this context, we observed that women of genetically exogenous provenance tend to be spatially integrated into their reproductive partner’s burial area, suggesting social integration into the host group (Fig. 1a). However, considering the 42 reproductive unions observed across all pedigrees that evidently had offspring buried at Gurgy, we noticed a shortage of mothers, with only 9 versus 20 fathers buried there. This imbalance is also observed in the total number of adult burials (38 male versus 20 female), and suggests that male adults were twice as likely to be buried than female adults. We therefore observed a potential sex bias in burials independent of female exogamy. This could be explained by different funerary practices being reserved for these mothers, or by other social factors mitigating against a co-burial with their reproductive partner’s group.
To gain independent information on individual mobility, we performed strontium isotope analyses (87Sr/86Sr) using laser ablation20 on 57 individuals (Fig. 3, Supplementary Note 11, Supplementary Table 23 and Extended Data Figs. 3 and 8). Unrelated female adults and some of those with no parents at the site show lower 87Sr/86Sr ratio values compared with male individuals from the same generation (Fig. 3 and Extended Data Fig. 3). Although the geological reference map does not enable us to infer a specific geographical origin, this finding provides further evidence that these female individuals grew up in different places before joining the Gurgy community (Supplementary Note 11 and Extended Data Fig. 8). Published stable isotope data (carbon, nitrogen, sulphur) measured on bones highlight a significant, sex-biased dietary division in adults21. On average, male individuals yielded higher δ13C and δ15N, and lower δ34S values, than female individuals, which could reflect a separation by sex, but could also signal female mobility (Supplementary Table 24). Genetic sex determination of subadults enabled us to confirm this difference also in childhood (P = 0.01019), which could be explained by a sex-related differential treatment at certain ages, determined by social rules (Supplementary Note 14 and Extended Data Fig. 5). Notably, the funerary practices in Gurgy show a shift at around 7–8 years of age, when the children are buried with different types of grave goods compared with younger ages, and another shift at around 15–16 years of age when they are associated with the same grave goods as adults, which could reflect local age stages or other social thresholds. This pattern has previously been observed at other Neolithic sites in the northern half of France22 (Supplementary Table 1).