|
|
Post by Admin on Sept 6, 2013 8:31:41 GMT
Recent studies have revealed that 2–3% of the genome of non-Africans might come from Neanderthals, suggesting a more complex scenario of modern human evolution than previously anticipated. In this paper, we use a model of admixture during a spatial expansion to study the hybridization of Neanderthals with modern humans during their spread out of Africa. We find that observed low levels of Neanderthal ancestry in Eurasians are compatible with a very low rate of interbreeding (<2%), potentially attributable to a very strong avoidance of interspecific matings, a low fitness of hybrids, or both. These results suggesting the presence of very effective barriers to gene flow between the two species are robust to uncertainties about the exact demography of the Paleolithic populations, and they are also found to be compatible with the observed lack of mtDNA introgression. Our model additionally suggests that similarly low levels of introgression in Europe and Asia may result from distinct admixture events having occurred beyond the Middle East, after the split of Europeans and Asians. This hypothesis could be tested because it predicts that different components of Neanderthal ancestry should be present in Europeans and in Asians. For instance, similar levels of Neanderthal introgression are observed in Europe and in Asia (1), which has been interpreted as evidence for a single and limited episode of admixture between Neanderthals and the ancestors of Eurasians some 50–60 kya (1). This interpretation implies either that there has been no subsequent admixture between modern humans and Neanderthals when the formers colonized Europe some 40 kya (10⇓–12) or that some admixture occurred in Europe, where these species coexisted (13), but that this signal has now disappeared because of drift (14) or later migrations of nonadmixed populations (1). Moreover, the very low level of Neanderthal ancestry observed in Eurasians (1.9–3.1%) (2) is somehow surprising, because one would expect to see massive levels of Neanderthal introgression into modern humans if admixture was not strongly prevented during the range expansion of modern humans out of Africa (3, 15).  Under our model of range expansion of modern humans competing locally with Neanderthals, admixture can only occur in a narrow zone on the expansion front where the two populations coexist and have the possibility of hybridizing (3). To have similar final rates of introgression, the number of hybridizations having occurred on the Asian wave front must be roughly similar to that the number of hybridizations having occurred on the European wave front, which explains why the restricted Neanderthal range often leads to unequal introgression rates in Europe and Asia. Our result are thus in line with a recent paleogenetic study demonstrating that some hominin remains in the Altai Mountains are from Neanderthal type (18) and that Neanderthal range thus extended further east than previously believed. Although we have modeled the Asian range to extend up to the Altai region north of the Himalayas, we cannot be certain that the ancestors of East Asians migrated through this region. However, the facts that Papua New Guineans show signals of hybridization with another hominin (Denisovan) (2) and that their ancestors are likely to have followed a coastal southern route to the Pacific (19, 20) suggest that the Denisovan range must have extended more to the south and that the ancestors of East Asians may have indeed traveled north of the Himalayas, above the Denisovan range. www.pnas.org/content/108/37/15129.full The modern human haplotypes that possess signals of introgression, including the microcephalin D-haplogroup (Evans et al., 2006) and the H2 haplotype of the MAPT locus (Hardy et al., 2005) as well as the set of Neanderthalspecific SNPs identified by Plagnol & Wall (2006) are in need of empirical testing and verification in multiple Neanderthal samples. Widespread distribution in Neanderthals of either the microcephalin D-haplogroup or the MAPT H2 haplotype would be highly indicative of successful interbreeding between modern humans and Neanderthals. webh01.ua.ac.be/funmorph/raoul/fylsyst/herrera2009.pdf |
|
|
|
Post by Admin on Sept 6, 2013 22:46:48 GMT
.PNG/300px-HG_J1_(ADN-Y).PNG) Tracing the origin and expansion of pastoral nomadism in the Middle East has widespread significance for understanding the development of the civilizations of the ancient Near East and the spread of the Semitic languages throughout the Levant, the Arabian Peninsula and Mesopotamia. Y-chromosome analyses of modern populations of the Middle East can contribute to the delineation of the demographic and migration processes in this region. The predominant categories of Y chromosomes in this region are varieties associated with haplogroup J-M304. The timing and geographical distribution of J1e is representative of a demic expansion of agriculturalists and herder–hunters from the Pre-Pottery Neolithic B to the late Neolithic era.24, 26 The higher variances observed in Oman, Yemen and Ethiopia suggest either sampling variability and/or demographic complexity associated with multiple founders and multiple migrations. The expansion time associated with Yemen is somewhat older (7000 BCE) and may reflect a migration of herders into southern Arabia.27 Finally, the more recent expansion times (Table 1) observed in Arabs from the Arabian Peninsula, Negev Bedouins and Sunni Arabs from Hama, Syria, are consistent with a subsequent Chalcolithic/Early Bronze Age (3000–5000 BCE) advance of J1e to the Arab populations of Arabia from near the early attested Arabian-speaking area of Tayma in north central Arabia28, 29 (Figure 1f).  Figure 2 Figure 2
(a) Median-joining network for J1* using the nine-locus Y-STR haplotypes. Networks were weighted according to Qamar et al.22 Loci analyzed included DYS19, DYS388, DYS389I, DYS389II, DYS390, DYS391, DYS392, DYS393 and DYS439. (b) Median-joining ...Although on a broad scale the haplogroup J1e frequency distribution and expansion times are consistent with the model that it tracks a possible expansion of Neolithic agro-pastoralists from the Fertile Crescent into the arid Arabian Peninsula, several caveats must be considered. First, the patchy distribution of J1e frequency in the Levant (Syria, Jordan, Israel and Palestine) may reflect the complex demographic dynamics of religion and ethnicity in the region. Second, even though the highest YSTR variance of J1e lineages is in eastern Anatolia, northern Iraq and northwest Iran, one cannot entirely rule out recent admixture as a contribution to the high variance among ethnic Assyrians. A recent Bayesian analysis of Semitic languages supports an origin in the Levant 5750 years ago and subsequent arrival in the Horn of Africa from Arabia 2800 years ago,11 thus providing an indirect support of our phylogenetic clock estimates. It is important to note that the glottochronological dates yield estimates for the break-up and expansion of the Proto-Semitic language. Proto-Semitic, itself, may have been spoken in a localized linguistic community for millennia before its bifurcation into the East and West Semitic branches. In summary, haplogroup J1e data suggest an advance of the Neolithic period agriculturalists/pastoralists into the arid regions of Arabia from the Fertile Crescent and support an association with a Semitic linguistic common denominator.14 www.ncbi.nlm.nih.gov/pmc/articles/PMC2987219/#!po=28.9474 |
|
|
|
Post by Admin on Sept 10, 2013 5:36:07 GMT
 MDS Plot of FST Genetic Distances Generated from mtDNA HVS1 Sequences in Siberian and Central Asian Populations MDS Plot of FST Genetic Distances Generated from mtDNA HVS1 Sequences in Siberian and Central Asian PopulationsThe Altai region of southern Siberia has played a critical role in the peopling of northern Asia as an entry point into Siberia and a possible homeland for ancestral Native Americans. It has an old and rich history because humans have inhabited this area since the Paleolithic. Today, the Altai region is home to numerous Turkic-speaking ethnic groups, which have been divided into northern and southern clusters based on linguistic, cultural, and anthropological traits. To untangle Altaian genetic histories, we analyzed mtDNA and Y chromosome variation in northern and southern Altaian populations. All mtDNAs were assayed by PCR-RFLP analysis and control region sequencing, and the nonrecombining portion of the Y chromosome was scored for more than 100 biallelic markers and 17 Y-STRs. Based on these data, we noted differences in the origin and population history of Altaian ethnic groups, with northern Altaians appearing more like Yeniseian, Ugric, and Samoyedic speakers to the north, and southern Altaians having greater affinities to other Turkic speaking populations of southern Siberia and Central Asia. Moreover, high-resolution analysis of Y chromosome haplogroup Q has allowed us to reshape the phylogeny of this branch, making connections between populations of the New World and Old World more apparent and demonstrating that southern Altaians and Native Americans share a recent common ancestor. These results greatly enhance our understanding of the peopling of Siberia and the Americas. europepmc.org/articles/PMC3276666/reload=0;jsessionid=Isv22mSDw88Jww2wG9Qp.4A study revealed that the ancient people, who established the Moche civilisation in the Lambayeque region in Peru from about 100 AD to 800 AD, had genetic links to the modern populations of Siberia and the Ainu people of northern Japan. It's also known that "Kennewick Man", the skeletal remains of a prehistoric man found in Kennewick, is genetically closer to the Ainu than Native American tribes and the proto-Japanese people reached America around 7300 BC to 7600 BC after crossing the Bering Strait.  The first three principal components from the principal components analysis of all 52 variables account for only 39.22% of the total size-corrected variation in the data. The PCA plot (Figure 3) places the Kennewick individual as an outlier compared to full Howells data (N = 22 populations). Using the principal component scores to generate inter-individual distances (Van Vark and Schaafsma 1991), the Kennewick individual is closest to south Pacific (Moriori, Easter Island) and the Ainu of Japan. The typicality probabilities for the PC reduced data, which are the least conservative estimates of group membership, all indicate that the Kennewick cranium is not morphologically similar to any modern human population (Table 7). No modern Native American group is included as a close neighbor in the least conservative approach, which strongly suggests that they bear no morphological resemblance to the Kennewick remains. Furthermore, while the inclusion of the Ainu as a nearest group could be interpreted as a possible "Caucasoid" morphology for the Kennewick remains if one considers modern Ainu to be "Caucasoids" (see Jantz and Owsley 1997); we view this as a reflection of the southern Asian/south Pacific morphology of the Kennewick skull given that most researchers tend to associate Ainu groups with earlier population originating in southern Asia (Brace and Hunt 1990; Turner 1985, 1990). Overall, the Native American mtDNA genomic data suggests the following scenario for the peopling of the Americas. The transition period between the separation of the Native American mtDNA haplogroups from their Asian ancestors and the start of their diversification and expansion into the Americas was estimated at > 5000 years. Adding this time to our estimates for the expansion indicates that the beginning of the divergence of the Native American founder population from its Asian ancestral population probably predates the LGM. There is evidence of human settlements in the artic around 30 kya.57 Therefore, it is possible that the precursors of Native American populations represent a human group that pioneered colonization of northeast Asia before the LGM, during a period of amelioration in the climatic conditions.58 Although it was not possible to determine where in northeast Asia this population stayed during this long period of isolation, Beringia represents the best candidate for that location, at least for the moderate bottleneck period (∼20 kya) before the expansion. Toward the end of the Pleistocene, Beringia was mostly exposed, and even though archaeological evidence for human presence in Beringia around the LGM is controversial, the first evidence of human settlements around this area predates the LGM.57 There is also strong evidence that the Beringian environment could sustain human populations during at least part of the LGM, being considered both climatically and ecologically a glacial refugium.59 mtDNA results further suggest that during the LGM the Native American founding population experienced a reduction in size (with a minimum of around 1000 women) that lasted for 3000–4000 yr and may have been linked to the deteriorating conditions of Beringia at the LGM.60 In addition, the Beringian refugium was ecologically isolated to the west and physically isolated to the east by the glaciers that seem to have effectively blocked the way to America until near the end of the LGM; this may help to explain why the population stayed in Beringia for such a long time before expanding south. We estimate that beginning ∼19–18 kya and ending ∼16–15 kya (i.e., toward the end of the LGM), the Native American founding population experienced a significant demographic growth process that is most likely associated with an extensive range expansion and may mark the beginning of the effective colonization of the New World south of Beringia. Given that the opening of the ice-free corridor is dated not earlier than ∼14 kya, our results strongly support an alternative route for this expansion, most likely along the western coast of North America.61,62 Recent data have shown that this coastal route was largely ice free by ∼19 kya and that the environment improved rapidly, being capable of supporting bears as of ∼15 kya.63 Interestingly, the end of the intense expansion period coincides with the age of the southern South American Monte Verde site, ∼14.5 kya.3 The strong and rapid population growth suggested by our data is consistent with a model in which humans have traveled the > 13,000 km along the coast from Alaska to the southern tip of Chile in a few thousand years.64 www.ncbi.nlm.nih.gov/pmc/articles/PMC2427228/ |
|
|
|
Post by Admin on Sept 13, 2013 16:19:08 GMT
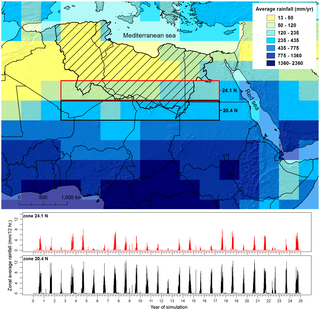 Figure 1. Palaeorainfall used to drive the combined numerical model. Figure 1. Palaeorainfall used to drive the combined numerical model.
Yearly average rainfall from a 25 year snapshot of an ESM experiment and catchment area (hatched region) as well as the time series of zonally averaged precipitation for two stripes south of the catchment highlighting the (i) South-North gradient of rainfall during the wet seasons (June to September) and (ii) the modelled year to year variability of the monsoon system.The role of the Sahara as a geographical filter and launch zone for dispersals of Homo sapiens out of Africa is a controversial topic [1], [2], [3], [4]. At issue is the observation that 130-100,000 years ago there was a marked increase in humidity in the present desert and adjacent regions [5], [6], [7], [8], which coincided with some of the earliest appearances of H. sapiens in both the Sahara and the Levant [9], [10], [11], [12], [13], [14], [15], [16]. During MIS 5 (Marine Isotope Stage 5) [17], high insolation in the northern hemisphere caused the African monsoon to assume a position up to 1,000 km north of its location today [18], [19], [20], [21]. Isotopic and geomorphic evidence suggests that rain falling on the north of the Trans-Saharan mountains then flowed towards the Mediterranean [2], [22], [23], potentially creating migration pathways via a series of ‘green corridors’ [4] and ‘mega lake corridors’ [24] across the Sahara. Dating of human fossils from the Levantine sites of Skhul and Qafzeh imply that early dispersals occurred along the eastern margins of the Sahara prior to ~100 ka [13], [14], [15], while other craniodental remains show that populations closely resembling those of the Near East were simultaneously present in north western Africa [9], [16], [18]. Given the combined dating uncertainties, many of these fossils and associated archaeological Middle Palaeolithic/Middle Stone Age finds are likely contemporary with the last interglacial period of peak humidity attested in marine cores [20], [23] and stalagmites [6], [25]. 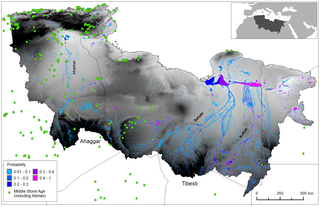 Figure 2. Simulated probability of surface water during the last interglacial. Figure 2. Simulated probability of surface water during the last interglacial.
This figure details Archaeological sites, and an annual probability that a location has surface water. The archaeological data are derived from a number of sources (including [42], [66], [67], [68]. The findspots are characterised by Aterian and Middle Stone Age artefacts such as bifacial foliates and stemmed Aterian points and/or typical ‘Mousterian’ points, side scrapers and Levallois technology.Figure 2 demonstrates the existence of a series of extensive ephemeral and perennial river systems draining North from the Ahaggar and Tibesti mountains across the Sahara to the Mediterranean during the period 125–124 ka BP. Some channels dissipate in the desert, but some converge forming three main systems; in the West the Irharhar river draining into the Chott Melrir basin, and to the East two larger systems named the Sahabi and the Kufrah (Fig. 2). As the rainfall is associated with the monsoon, flow is highly seasonal and the Irharhar river is ephemeral, flowing for ~3 months. The Sahabi and Kufrah systems are close to perennial, due to larger contributing areas in the catchment headwaters located in the monsoon belt (Fig. 1). 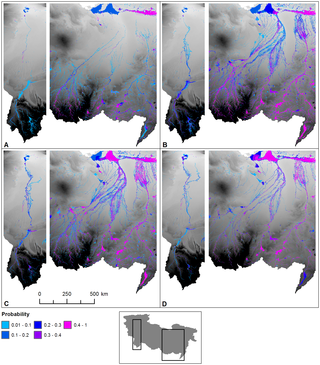 Figure 3. Seasonal flow of surface water across the Sahara during the last interglacial. Figure 3. Seasonal flow of surface water across the Sahara during the last interglacial.
Monthly probability of surface water being present for (a) August, (b) September, (c) October and (d) November. These illustrate the hydrodynamics of the system simulated by the large scale 2d hydraulic model that routes a flood wave of water north across the desert from the mountains.Figure 3 demonstrates the seasonality of flow in all three systems with precipitation in August taking over two months to reach the coast or near-coastal lake systems. The distinct climatic zoning (Fig. 1) means all three rivers are allogenic, losing water along their length with little or no hydrological contribution once they leave the mountains/uplands. In addition to rivers, the simulations predict massive lagoons and wetlands in NE Libya some of which are extensive (>70,000 km2). These are also fed from the Jebel Akhdar in Cyrenaica, which also received higher rainfall during this period. There are smaller lakes forming in Tunisia and Algeria due to water supplied via the Irharhar river system. This study provides the first strong quantitative evidence for the presence of three major river systems flowing across the Sahara during MIS 5e. We simulated three river systems that are now largely buried by dune systems, but when flowing would have provided fertile habitats for flora and fauna in proximity to the channels [22]. Notably, the Sahabi and Kufrah would be major river systems with monsoon discharges significantly in excess of 2500 m3 s−1 and an extensive system of anabranches and wetlands. In the Libyan Kalanschiou region, the green corridor would have been 100 km wide, substantial and largely perennial. This reconstruction is highly compatible with evidence of widespread palaeosols deposited on the margins of this system during the less pronounced Holocene humid period [22]. Here we have simulated one wet phase, but this research strongly supports the occurrence of similar ‘Green Saharas’ recorded in the marine [37] and terrestrial [26] archive. www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0074834 |
|
|
|
Post by Admin on Sept 16, 2013 21:33:51 GMT
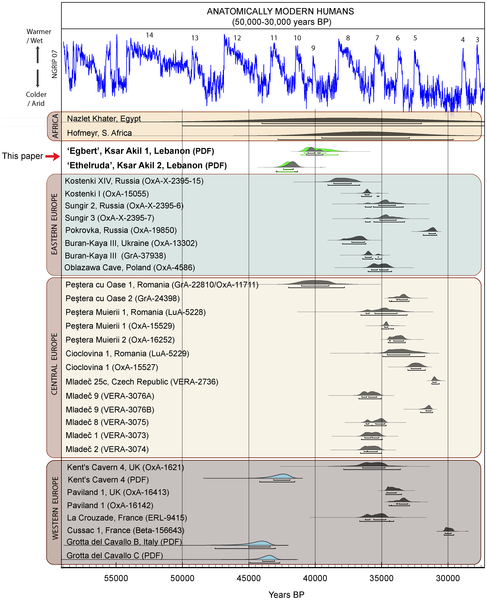 Comparison of the modelled ages (Probability Distribution Function; PDF) obtained for Egbert and Ethelruda with age estimates of AMH from other Palaeolithic sites between 50,000–30,000 years ago. Comparison of the modelled ages (Probability Distribution Function; PDF) obtained for Egbert and Ethelruda with age estimates of AMH from other Palaeolithic sites between 50,000–30,000 years ago. Levant is the most likely place where admixture events between Levantine Neanderthals and early Homo sapiens took place and the two species of ancient humans occupied the region interchangeably from the Upper Paleolithic to the Middle Paleolithic, creating the opportunity to a certain number of successful interbreeding events leading to the 1 to 4% introgression of Neanderthals into Eurasians. There are skeletal morphological similarities among European late Neanderthals and early Homo sapiens fossils that hint at such interbreeding after 45 ka and Levantine Homo sapiens and Neanderthals are less morphologically distinct from one another than their European counterparts (Zilhao, 2006). Despite increasing archaeological and genetic data in support of an African origin for modern humans, there is little consensus about the exact timing or about the route or routes taken during migration(s) out of Africa and into Asia and Europe [58]. The notion that modern humans dispersed first into the Near East and then directly into Europe, is a common perception amongst palaeoanthropologists and prehistorians. It is currently accepted that modern humans migrated from Africa in several waves, probably using a number of dispersal routes. Garcea [59] distinguishes two Out of Africa movements by AMH, on the basis of individual features and of being separated by a long time span. The earlier wave or “Out of Africa 2a” took place between about 130,000 and 80,000 years ago, while the second “Out of Africa 2b” occurred at ~50,000 years ago, after an apparent gap of about 30,000 years [59], [60]. While the Levant appears an obvious route in and out of Africa based solely on its geographical position, there is, as yet, no evidence for significant human and animal migrations during the Pleistocene ([67], and papers therein), let alone during the short time window of the Middle to Upper Palaeolithic transition or during the “Out of Africa 2b” scenario. The current evidence for contemporaneity in the appearance of both transitional/IUP and EUP technocomplexes in Europe and the Levant implies that the northern Mediterranean Levantine coast might not be the point of origin for the dispersal of the earliest Upper Palaeolithic outwards and into Europe. This, in turn, suggests to us that current models based on old assumptions regarding the pathway(s) of human dispersal require further testing and, possibly, revision. The Levantine late Middle Palaeolithic is solely associated with Neanderthals [68], and Ewing's attribution of Ethelruda to a “Neandertaloid” individual has helped shaping a generalized view that Neanderthals may have been involved in the making of the transitional/IUP layers, at least in its earliest phases (e.g. [5]). The most recent archaeological and palaeoanthropological data from Ksar Akil with regards Ethelruda (e.g. [35]), as well as the likely-modern teeth from Üçağızlı, where Mousterian levels have not been identified, may provide a contrary view to traditional assumptions on the authorship of the IUP by Neanderthals. Of course, it is wise to remain cautious on the taxonomic status of both Ethelruda and the teeth from Üçağızlı until more detailed scientific work has been undertaken using state-of the-art methodologies. Further work is also urgently required to address whether other technocomplexes exhibiting similar characteristics, such as the Balkan Bachokirian (e.g. level 11 of the eponymous site, containing fossil remains; [69]) or the Bohunician (or Emiro-Bohunician; [70]) of Central Europe, fall into the same category. www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0072931 Fig. 2. Locations of the principal dated Early Upper Palaeolithic archaeological sites discussed in this paper. Fig. 2. Locations of the principal dated Early Upper Palaeolithic archaeological sites discussed in this paper.Around 80–75 ka, global temperatures dropped precipitously. Rampino and Self (1992) have implicated the eruption of the Mt. Toba (Indonesia) and a subsequent long-lasting ‘‘volcanic winter’’ in this climate change. At this point in time, the Levant appears to have become abruptly colder and drier, more like Central Asia Homo sapiens populations already living in the Levant were probably increasingly confined to perennial water sources and refugia of woodland vegetation along the Central and Northern Levant coast. The symbolic evidence we see at Skhul and Qafzeh, the use of personal adornments, production of mineral pigments, and mortuary furniture, may reflect efforts to cope with the social stresses of population packing by increased use of exosomaticsymbolic media in social interactions. Neanderthals reoccupied the Levant when warmer and more humid conditions reasserted themselves after 70 ka. The archaeological record is not clear whether Late MP Neanderthal adaptive strategies were substantially different from those of their Homo sapiens precursors at this point in time. If they were not, then it is possible that Neanderthal success simply reflects a greater physiological tolerance for colder conditions (a quality one might reasonably infer for a species that evolved on the cold, dry steppes of western Eurasia) (Aiello and Wheeler, 2003). Why the Levant was not also rapidly reoccupied by African humans remains unknown. There are hints from recent human genetic variability of substantial reductions in African human populations around this period (Ambrose, 2003). It is possible that population densities among Homo sapiens populations living in those parts of Africa nearest to the Levant were so low that there was no strong selective pressure to disperse northward into the Levant until tens of millennia later. Alternatively, human dispersal out of Africa may have followed a different, southern route, one bypassing the wellentrenched Levantine Neanderthals by expansion into southern Asia along the coast of the Indian Ocean (Kingdon, 1993; Lahr and Foley, 1994; Macaulay et al., 2005; Beyin, 2006).The period ca 50–45 ka in the Levant also witnessed significant short-term cooling correlated with the North Atlantic Heinrich H5 Event. What appears to have been an impressively widespread Neanderthal occupation probably retracted towards the Mediterranean coast. Late MP sites located in the coastal lowland Levant provide numerous indications of increased occupational ‘‘intensity’’ (Lieberman and Shea, 1994, p. 318), that is, either more prolonged occupations or shorter intervals of site abandonment and reoccupation. This evidence includes more substantial hearths (Meignen et al., 2005), increased percentages of less-desirable large mammal prey (Speth and Clark, 2006), multiseasonal site occupations (Lieberman, 1998), increased use of specialized hunting equipment (Shea, 1995), increased use of expedient core technology (Wallace and Shea, 2006), and increased use of hard-seeded plant foods (Madella et al., 2002; Lev et al., 2005). Late MP hominins appear to have been working harder and harder to maintain themselves than their predecessors. In spite of this effort, there does not appear to have been any substantial increase in Neanderthal geographic range beyond the Levant to Mediterranean North Africa or the Nile Valley. Nor does this intensification seem to have led to long-lasting changes in Neanderthal strategic ecology. Instead, Neanderthal fossils cease to appear in the Levantine fossil record after 45 ka, an observation exactly consistent with their inferred extinction. Much as is the case elsewhere in western Eurasia, there is no clear and convincing evidence for overlapping Neanderthal and Homo sapiens occupations.  Fig. 5. Changes in Levantine climate 130–25 ka as reflected in Soreq and Peqiin Speleothem Data (graphic generated using data from Bar-Matthews et al., 2003b). The broken line indicates approximate modern values for δ18O. Fig. 5. Changes in Levantine climate 130–25 ka as reflected in Soreq and Peqiin Speleothem Data (graphic generated using data from Bar-Matthews et al., 2003b). The broken line indicates approximate modern values for δ18O.Levantine Homo sapiens and Neanderthals are less morphologically distinct from one another than their European counterparts (Kramer et al., 2001; but see Collard and Franchino, 2002), nevertheless, clear and convincing proof of gene flow between Levantine Neanderthals and Homo sapiens remains elusive. Deterioration of DNA in fossils from warm climates makes it unlikely that palaeo-genetics will resolve this issue for the Levant (Smith et al., 2003). Clearly, this is a situation that calls for multiple working hypotheses, not just continuing to assume Neanderthals and Homo sapiens interbred whenever possible. What obstacles might have existed for interbreeding between Neanderthals and Homo sapiens? Even if one sets aside the stratigraphic problems with evidence for Neanderthal–Homo sapiens sympatry, there is an ecological objection to this hypothesis. The similarities we see among Levantine Neanderthals and Homo sapiens settlement patterns, subsistence strategies, and technological organization suggest they occupied essentially identical ecological niches (Shea, 2003a,b). Much of what we know about Eurasian MP and African MSA subsistence suggests these niches were similar to those of recent large social carnivores (Stiner, 1993; Marean and Assefa, 1999). Among large carnivores, intra- and interspecific competitive encounters over resources are significant sources of mortality (Van Valkenburgh, 2001). Even if they might have been able to interbreed, Neanderthal and Homo sapiens populations living in a small geographically circumscribed region like the Levant had powerful evolutionary incentives for avoiding each other. Such competition may have been less intense (and opportunities for interbreeding greater) in less circumscribed regions, but this hypothesis needs to be refined with palaeoanthropological evidence from other regions and studies of hybridizing behaviour among living mammals (Holliday, 2003). 6.2. Predictions of the turnover hypothesis If rapid climate change in the Levant ca 75 ka and 45 ka led to extinctions and turnovers in the region’s hominin populations, it follows that both prior and subsequent episodes of rapid climate change should have had similar effects. Evidence from the earliest and latest phases of the Late Pleistocene in the Levant supports this hypothesis. Transitions or turnovers? Climatically-forced extinctions of Homo sapiens and Neanderthals in the East Mediterranean Levant |
|

