|
|
Post by Admin on Sept 21, 2013 20:49:18 GMT
Europeans and Asians have inherited 1-5% of Neanderthal DNA as Homo sapiens out of Africa interbred with the Neanderthals in the Mediterranean around 60,000 years ago, which gave rise to the IQ gap between non-Africans and Africans with no Neanderthal admixture. Neanderthal cranial capacity was slightly larger than that of modern humans and they invented stone tools known as 'flakes', which were more efficient than the blades adopted by Homo sapiens, and the Neanderthals had more sophisticated Stone Age culture than AMHs who had just reached Europe. Previously, advanced bone tools have only been associated with modern humans but the following study (Soressi et al. 2013) puts these pieces in proper Neanderthal contexts, concluding that the Neanderthals may have had a civilising influence over Upper Paleolithic modern human populations in Europe where lissoirs were commonly used. Modern humans replaced Neandertals ∼40,000 y ago. Close to the time of replacement, Neandertals show behaviors similar to those of the modern humans arriving into Europe, including the use of specialized bone tools, body ornaments, and small blades. It is highly debated whether these modern behaviors developed before or as a result of contact with modern humans. Here we report the identification of a type of specialized bone tool, lissoir, previously only associated with modern humans. The microwear preserved on one of these lissoir is consistent with the use of lissoir in modern times to obtain supple, lustrous, and more impermeable hides. These tools are from a Neandertal context proceeding the replacement period and are the oldest specialized bone tools in Europe. As such, they are either a demonstration of independent invention by Neandertals or an indication that modern humans started influencing European Neandertals much earlier than previously believed. Because these finds clearly predate the oldest known age for the use of similar objects in Europe by anatomically modern humans, they could also be evidence for cultural diffusion from Neandertals to modern humans. Specialized bone technology first appears in Africa (1⇓⇓⇓–5) and is widespread in Europe after the arrival of modern humans with the beginning of the Upper Paleolithic (6⇓–8). Modern humans shaped bone by grinding and polishing to produce standardized or so-called formal tools that were used for specific functions (6, 9). Examples of Neandertal bone tools do exist (9⇓⇓⇓⇓⇓–15); however, most of these were made through percussion to mimic existing stone tools, such as handaxes, scrapers, and denticulates. Standardized bone tools in forms distinct from stone tools and shaped by grinding and polishing occur in the Châtelperronian (16) and Uluzzian (17), but (i) whether Neandertals made these assemblage types is debated and, furthermore, (ii) their late date means that Neandertals could have been influenced by modern humans already in Europe (18, 19). Examples with earlier dates are disputed (20). For example, the site of Saltzgitter-Lebenstedt yielded several mammoth ribs modified by percussion and then shaped by grinding (12).  Fig. 3.Photomicrographs of the Pech I (G8-1417) bone showing details of the polish and striations (A–D). Use-wear traces on the upper side of an experimental bone lissoir used to soften dry hide with a longitudinal motion after 5 min of use (E) and after 10 min of use (F). Fig. 3.Photomicrographs of the Pech I (G8-1417) bone showing details of the polish and striations (A–D). Use-wear traces on the upper side of an experimental bone lissoir used to soften dry hide with a longitudinal motion after 5 min of use (E) and after 10 min of use (F). Fig. 4. Examples of Upper Paleolithic lissoirs. Distal fragment of a Gravettian lissoir from Abri Pataud (France) (45) (A). Distal fragment of a Proto-Aurignacian lissoir from La Grotte du Renne (France) (42) (B) and of a Magdalenian lissoir from La Grotte de la Vache (France) (24) (C). Almost complete Aurignacian lissoir from Gatzarria (France) (D) and mesial fragment (with typical scars of bending fractures at both ends) of a Aurignacian lissoir from Castanet-Nord (France) (46) (E). An unused modern lissoir (upper end) and plior (bottom end) use by leather craftsmen and made from a cow rib, purchased from the Internet, January 2013 (F). [(A) Collection MNHN, photo by C. Vercoutère; (C) Photo by E. Tartar; (D) Castanet project archives.] Fig. 4. Examples of Upper Paleolithic lissoirs. Distal fragment of a Gravettian lissoir from Abri Pataud (France) (45) (A). Distal fragment of a Proto-Aurignacian lissoir from La Grotte du Renne (France) (42) (B) and of a Magdalenian lissoir from La Grotte de la Vache (France) (24) (C). Almost complete Aurignacian lissoir from Gatzarria (France) (D) and mesial fragment (with typical scars of bending fractures at both ends) of a Aurignacian lissoir from Castanet-Nord (France) (46) (E). An unused modern lissoir (upper end) and plior (bottom end) use by leather craftsmen and made from a cow rib, purchased from the Internet, January 2013 (F). [(A) Collection MNHN, photo by C. Vercoutère; (C) Photo by E. Tartar; (D) Castanet project archives.]These bone tools are identical in outline, profile, and use-wear to lissoirs (Fig. 4 and SI Appendix, Section S6). Lissoirs are known from the early Upper Paleolithic of western Europe (6), including the Châtelperronian (16, 41), Proto-Aurignacian (42), and Aurignacian (22, 23), but are also found in the late Upper Paleolithic (24) through to historic and modern time periods (Fig. 4E). Lissoirs have a standardized shape and vary in size depending on the species used; they are an effective tool for producing and smoothly shifting pressure over a small area (21). This technique, when applied to animal skins, results in tougher, more impermeable, and lustrous hides (21). No other known artifact in the Middle or Upper Paleolithic toolkit could accomplish this task, meaning that these tools exploit specific properties of bone for shaping and use (21). king The bones reported here demonstrate that Middle Paleolithic Neandertals were shaping animal ribs to a desired, utilitarian form and, thus, were intentionally producing standardized (or formal) bone tools using techniques specific to working bone. These bones are the earliest evidence of this behavior associated with Neandertals, and they move the debate over whether Neandertals independently invented aspects of modern human culture to before the time of population replacement. In central Europe, artifact assemblages contemporaneous with the Pech I Neandertals but more comparable to assemblages from southwest Asia made by modern humans (19, 43) have such poor bone preservation that neither human fossils nor bone tools are known, and thus their influence on Neandertals cannot be evaluated. Thus, it remains to be determined whether MTA lissoirs are evidence that modern humans influenced Neandertals earlier and longer than previously suggested, whether these lissoirs represent independent invention and convergence, or whether, perhaps this time, Neandertals may have influenced subsequent Upper Paleolithic modern human populations in western Europe where lissoirs are common. www.pnas.org/content/110/35/14186.full |
|
|
|
Post by Admin on Oct 1, 2013 20:53:36 GMT
 Mitochondrial DNA from Neanderthals has not been found in humans and genetic evidence suggests that male humans and female Neanderthals did not mate. Human-Neanderthal admixture was extremely rare at the time and it is likely to have taken place in some forms of rape by Neanderthal males who encountered female H. sapiens on rare occasions and their offspring contributed to spreading Neanderthal DNA among AMH. Male humans may not have found female Neanderthals attractive or female Neanderthals were protected by stronger Neanderthal males, making them unavailable for male H. sapiens. Studies have shown though that modern humans share a common male ancestor who lived 59,000 years ago. Could this male ancestor have been Neanderthal? Indeed, the date of our closest common male ancestor correlates well with estimations of gene flow between Neanderthals and humans around 50,000 to 80,000 years ago. If H.neanderthalensis and H.sapiens were able to mate, then it is plausible that only the male H.neanderthalensis and the female H.sapiens were able to produce fertile offspring. The reciprocal cross may not have been successful. According to Haldane’s law, the heterogametic offspring of interspecific hybrids are likely to be absent, rare, or sterile (Short, 1997). If Haldane’s Law applied to the offspring of H.neanderthalensis and H.sapiens, we would expect to find female hybrids quite commonly, but male hybrids much more rarely. The male hybrids would have carried a Y chromosome very similar to that of the original hybridizing male. The lack of Neanderthal mtDNA suggests that the initial hybridization involved a Neanderthal male, but there would probably have been few if any male hybrid offspring, so the Neanderthal Y chromosome and the mtDNA would have been quickly lost. Nonetheless, the Neanderthal autosomes would have happily mingled and interchanged with human autosomes, eventually losing their identity in the process. Could it be that Homo neanderthalensis males were able to mate with Homo sapiens females but that the reciprocal cross was unsuccessful? Alternatively, were male H.sapiens disastrously incapable of wooing the physically more powerful H.neanderthalensis females? Or were H.neanderthalensis females simply unable to give birth to hybrid offspring? Perhaps male H.neanderthalensis outcompeted early male H.sapiens and eventually the male Neanderthal genes gained dominance (and maybe H.sapiens females somehow out-competed H.neanderthalensis females for partners). All of these possibilities potentially explain how we share a common male ancestor 59,000 years ago, but a common female ancestor 170,000 years ago. Simultaneously, these hypotheses explain why comparisons of DNA sequences in mitochondrial DNA from Neanderthals and modern humans have indicated that there was no interbreeding between these two exceedingly similar species (Potts & Short, 1999:59). Mitochondrial DNA from Neanderthals simply may not have made it into the modern human lineage. The nuclear DNA of Neanderthal males, however, possibly did. blogs.plos.org/neuroanthropology/2010/10/26/the-neanderthal-romeo-and-human-juliet-hypothesis/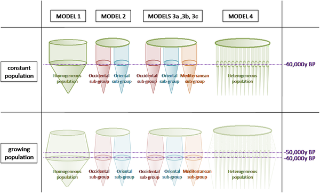 Figure 1. Schematic presentation of the six models. Figure 1. Schematic presentation of the six models.The third models correspond to an ancestral population which gives rise to three sub-populations: one in the West, another in the East and one in the South (fig. 1, table 1). This southern population corresponds to the paleoanthropological hypothesis concerning the presence of a Southern population [3], [15]–[18]. According to the geographical barriers and morphological evidence, we have established three different divisions. The fossil of El Sidron from a paleogeographic standpoint is closer to French fossils than to Italian and Croatian fossils. On the basis of morphological data it might be closer to the southern fossils (model 3b). Due to its geographical position, the fossil of Mezmaïskaya, discovered in the Caucasus, might be placed either in the eastern (model 3a) or in the western group (model 3b and 3c). These divisions are shown in table 2. For model 3 (a, b, c) we made the same assumptions as in model 2 regarding population growth, migration, population sizes, and generation time. Forty eight simulations of this model have been tested, sixteen by grouping. Most measures of genetic diversity fit the observed measures more closely than in the previous models. Indeed, if we consider a growing population in which migration occurs, we see plausible and best values of P(Co|C) for all models (3a, 3b and 3c) for simulation sets with twelve or nine sequences. The most precise fit is that of model 3c, which presents values of P(Co|C) closest to 0.5 (table 1). If we consider a growing population with no migration, only model 3c presents the best values of P(Co|C). Thus models three, which posit three groups among Neanderthals, and assume a growing population, seem to be most realistic, and model 3c is the most plausible one. 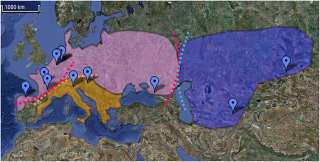 Figure 2. Map representing Neanderthal geographical distribution in groups. Models three and particularly model 3c are also statistically the strongest models. Moreover, they are coherent with the paleoanthropological data. Neanderthal features favour a distinction between western and eastern Neanderthals, with the western populations presenting more derived features than the eastern groups [13], [14]. This distinction has been explained by a migration to the East during the isotopic stages 5 or 6 [12], [14] or by a phenomenon like isolation through distance [40]. The sequence of Mezmaïskaya (in the Caucasus) is thus located in an intermediate region and could be placed in either of these groups. According to the geographical position of this fossil, this sequence could also belong to still another group. In the state of our knowledge, however, we are unable to test this assumption, since we need at least two sequences to establish a group. These models, 3a, 3b and 3c, also support previous paleoanthropological studies of the phylogenetic position of the fossil remains of Okladnikov in Siberia [26]. This fossil has been identified as a Neanderthal on the basis of its nucleotide sequence. Our results situate this fossil in the eastern Neanderthal group. Our results clearly favour the existence of one southern group which is present throughout model three. Our models 3b and 3c differ in regard to the position of El Sidron (northern Spain). According the model 3b the fossil of El Sidron is located in the southern group, wheras the model 3c situates it in the group extending from Spain to Belgium. In addition, we supposed that sequences of Monte Lessini in Italy and Vindija in Croatia allow to the same group because it exists a morphological closeness. This can be explained in terms of eco-geographical factors. Indeed these two regions are isolated in the north by the Alps and, in the range of datings that we consider for these sequences, a marine regression offers an easier passage between northern Italy and Croatia. www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0005151 |
|
|
|
Post by Admin on Oct 9, 2013 23:30:05 GMT
 Haplogroup J can be found at the highest frequency in the Near East (12%) followed by Europe (11%) but J1c and J1b1a1 are exclusively found in Europe and the authors argue that about 90% of the Ashkenazi J lineages have a European origin (Costa et al. 2013). Almost half of mtDNAs in west/central European Ashkenazi Jews belong to haplogroup K, declining to ~15% in east European Jews, and haplogroup K was also found in a significant group of Palestinian Arabs and it reaches a level of 17% in Kurdistan, indicating its Middle Eastern origin. The K1a1b1 and K1a9 subclades specific to Ashkenazi K lineages are dated to ~2.3 ka and it's plausible that these subclades mutated locally after the Jewish Diaspora populations of Middle Eastern origin migrated to Europe around 2,000 years ago. The fourth major Ashkenazi founder mtDNA falls within haplogroup N1b, which is found in Middle East, Egypt, Caucasus and Europe. Besides the four haplogroup K and N1b founders, the major haplogroup in Ashkenazi Jews is haplogroup H, at 23% of Ashkenazi lineages. Haplogroup H is a major European haplogroup (40–50%), which may point to the assimilation of mtDNAs indigenous to Europe through conversion and it can be concluded that around a quarter of the Ashkenazi founder mtDNAs originated in Europe.  The extent to which Ashkenazi Jewry trace their ancestry to the Levant or to Europe is a long-standing question5, which remains highly controversial3, 4, 6, 12, 13, 14, 16, 17. Our results, primarily from the detailed analysis of the four major haplogroup K and N1b founders, but corroborated with the remaining Ashkenazi mtDNAs, suggest that most Ashkenazi maternal lineages trace their ancestry to prehistoric Europe.  Previous researchers proposed a Levantine origin for the three Ashkenazi K founders from several indirect lines of evidence: shared ancestry with non-Ashkenazi Jews, shared recent ancestry with Mediterranean samples, and their absence from amongst non-Jews2, and this suggestion has been widely accepted4. However, our much more detailed analyses show that two of the major Ashkenazi haplogroup K lineages, K1a1b1a and K2a2a1 have a deep European ancestry, tracing back at least as far as the early and mid-Holocene respectively. They both belong to ancient European clades (K1a1b1 and K2) that include primarily European mtDNAs, to the virtual exclusion of any from the Near East. Despite some uncertainty in its ancestral branching relationships, a European ancestry seems likely for the third founder clade, K1a9. The heavy concentration of Near Eastern haplogroup K lineages within particular, distinct subclades of the tree, and indeed the lack of haplogroup K lineages in Samaritans, who might be expected to have shared an ancestral gene pool with ancient Israelites, both strongly imply that we are unlikely to have missed a hitherto undetected Levantine ‘reservoir’ of haplogroup K variation (Supplementary Note 1).  Furthermore, our results suggest that N1b2, for which a Near Eastern ancestry was proposed (with much greater confidence than for K) by Behar et al.2, is more likely to have been assimilated into the ancestors of the Ashkenazi in the north Mediterranean. Finally, our cross-comparison of control-region and mitogenome databases shows that the great majority of the remaining ~60% of Ashkenazi lineages, belonging to haplogroups H, J, T, HV0, U4/U5, I, W and M1 also have a predominantly European ancestry. The Ashkenazim therefore resemble Jewish communities in Eastern Africa and India, and possibly also others across the Near East, Caucasus and Central Asia, which also carry a substantial fraction of maternal lineages from their ‘host’ communities11, 25. Despite widely differing interpretations of autosomal data, these results in fact fit well with genome-wide studies, which imply a significant European component, with particularly close relationships to Italians3, 4, 6, 7. As might be expected from the autosomal picture, Y-chromosome studies generally show the opposite trend to mtDNA (with a predominantly Near Eastern source) with the exception of the large fraction of European ancestry seen in Ashkenazi Levites22. www.nature.com/ncomms/2013/131008/ncomms3543/full/ncomms3543.html |
|
|
|
Post by Admin on Oct 17, 2013 6:08:19 GMT
 This Wednesday, Oct. 9, 2013 photo shows the stone age recycling site cave at the north of Israel next to the city of Zichron Yaakov. From caves in Spain and North Africa to sites in Italy and Israel, archaeologists have been finding such recycled tools in recent years. The conference, titled " The Origins of Recycling," gathered nearly 50 scholars from about 10 countries to compare notes and figure out what the phenomenon meant for our ancestors. Recycling was widespread not only among early humans but among our evolutionary predecessors such as Homo erectus, Neanderthals and other species of hominids that have not yet even been named, Barkai said. Avi Gopher, a Tel Aviv University archaeologist, said the early appearance of recycling highlights its role as a basic survival strategy. While they may not have been driven by concerns over pollution and the environment, hominids shared some of our motivations, he said. "Why do we recycle plastic? To conserve energy and raw materials," Gopher said. "In the same way, if you recycled flint you didn't have to go all the way to the quarry to get more, so you conserved your energy and saved on the material."  At Qesem cave, a site near Tel Aviv dating back to between 200,000 and 420,000 years ago, Gopher and Barkai uncovered flint chips that had been reshaped into small blades to cut meat—a primitive form of cutlery. Some 10 percent of the tools found at the site were recycled in some way, Gopher said. "It was not an occasional behavior; it was part of the way they did things, part of their way of life," he said. He said scientists have various ways to determine if a tool was recycled. They can find direct evidence of retouching and reuse, or they can look at the object's patina—a progressive discoloration that occurs once stone is exposed to the elements. Differences in the patina indicate that a fresh layer of material was exposed hundreds or thousands of years after the tool's first incarnation. Participants in the conference plan to submit papers to be published next year in a special volume of Quaternary International, a peer-reviewed journal focusing on the study of the last 2.6 million years of Earth's history. Norm Catto, the journal's editor in chief and a geography professor at Memorial University in St John's, Canada, said that while prehistoric recycling had come up in past studies, this was the first time experts met to discuss the issue in such depth. Catto, who was not at the conference, said in an email that studying prehistoric recycling could give clues on trading links and how much time people spent at one site.  Recycling bones in the Middle Pleistocene: the early cases of Gran Dolina TD10-1 (Spain), Bolomor Cave (Spain) and Qesem Cave (Israel). Jordi Rosell, Ruth Blasco et. al. Area de Prehistoria, Universitat Rovira i Virgili (URV), Tarragona, Spain and IPHES, Institut Català de Paleoecologia Humana i Evolució Social, Tarragona, Spain. Archaeologists have used different kinds of data to identify recycling. Most approaches to recycling are based on lithic artifact attributes, and especially on surface alterations, that suggest a period of discard between different activity events. But, recycling can be also approached by means of faunal remains from bone damage. The bone breakage processes generate a high number of small and large-sized fragments, which are usually discarded. However, some of these rejected elements can show suitable morphological characteristics to be used for different purposes. On this basis, it is necessary to distinguish between the use of bone raw material from pre-existent carcasses of elephants (and other large animals) and recycling of fragments resulting from the extraction of marrow contained in bones. In the first case, bones are considered as raw material similar to lithic industry. In the second one, bones are not fractured with a technological purpose, but they are selected to be used in a subsequent process. Here we present some early cases of recycled bones from the Middle Pleistocene sites of Gran Dolina TD10-1 and Bolomor Cave in Spain and Qesem Cave in Israel. The studied elements seem to have been part of a previous faunal processing sequence (with nutritional objective) and later, they seem to have been selected among rejected items to be used or modified intentionally. These fragments are dated to MIS 9 and show bone damage produced by use (retouched and unmodified soft hammers) or present configured forms (bone artifacts). With this study, we try to provide data on the recycling activities on faunal remains in the Middle Pleistocene and discuss on the origin of this behaviour. media.wix.com/ugd//16f7f1_12374fa2b4be5bccc8a33ff113b0ed67.pdf |
|
|
|
Post by Admin on Oct 18, 2013 15:38:41 GMT
 In their report, the researchers – led by Georgian anthropologist David Lordkipanidze – write that it is "the world's first completely preserved hominid skull." And what a skull it is. It suggests that the earliest known members of the Homo genus (H. habilis, H. rudolfensis and H. erectus) may not have been distinct, coexisting species, after all. Instead, they may have been part of a single, evolving lineage that eventually gave rise to modern humans. Discovered alongside the remains of four other early human ancestors, all of which date to roughly 1.8-million years ago, "Skull 5," as the specimen is called has a relatively tiny brain case and a protruding lower jaw – anatomical features reminiscent of Australopithecus, a more primitive human ancestor. And yet, its upper jaw resembles that of a 2.3-million-year-old specimen from Ethiopia, thought to have belonged to H. habilis; its bulky browline bears a strong resemblance to that of H. erectus; and its long, vertically oriented upper face and the overall shape of its braincase are unmistakably Homo. An anatomical chimera, Skull 5 possesses a surprising combination of features that has never before been observed together in an early Homo fossil.  The site of Dmanisi, Georgia, has yielded an impressive sample of hominid cranial and postcranial remains, documenting the presence of Homo outside Africa around 1.8 million years ago. Here we report on a new cranium from Dmanisi (D4500) that, together with its mandible (D2600), represents the world's first completely preserved adult hominid skull from the early Pleistocene. D4500/D2600 combines a small braincase (546 cubic centimeters) with a large prognathic face and exhibits close morphological affinities with the earliest known Homo fossils from Africa. The Dmanisi sample, which now comprises five crania, provides direct evidence for wide morphological variation within and among early Homo paleodemes. This implies the existence of a single evolving lineage of early Homo, with phylogeographic continuity across continents. In the past two decades, excavations at the archaeological site at Dmanisi, Georgia, have revealed hominin fossils from the earliest Pleistocene, soon after the genus Homo first dispersed beyond Africa. Lordkipanidze et al. (p. 326; see the cover) now describe a fossil cranium from the site. Combined with mandibular remains that had been found earlier, this find completes the first entire hominin skull from this period.  Some fossils previously discovered at Dmanisi seemed to have links to H. erectus. But when the big lower jaw [of Skull 5] was found in 2000, some researchers suggested it belonged to a new species they called Homo georgicus. With the discovery of the new, fifth skull the researchers had to confront head-on the variation among all five. Age and sex probably account for much of it: The skulls are thought to have belonged to an elderly toothless male, two mature males, a young female, and an adolescent of unknown sex. This broad sample from one place and a short span of time is what makes Dmanisi an “exceptional site”. By analyzing the skull shapes with 3D computer-based methods, the researchers found that the range of variation in the group at Dmanisi was no greater than within living humans or chimps. The team concluded that all five skulls belong to a single, variable species. In the end, the team settled on the cumbersome moniker of Homo erectus ergaster georgicus, which recognizes the skull as an earlier Georgian form of H. erectus. |
|

