|
|
Post by Admin on Mar 31, 2020 21:15:14 GMT
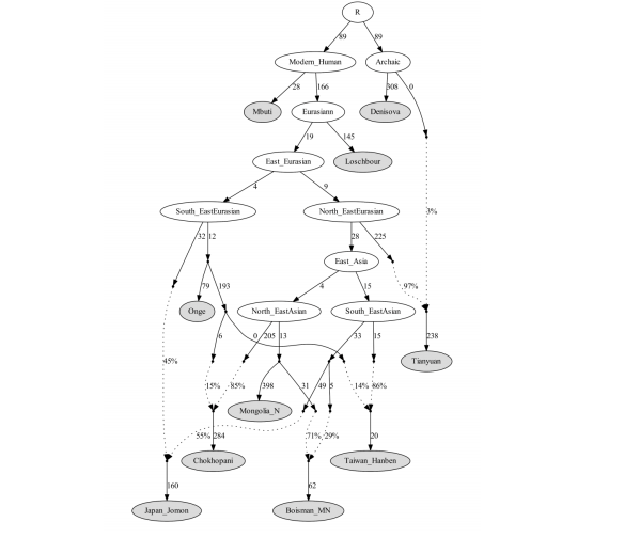 Figure 5: qpGraph modeling of a subset of East Asians. We used all available sites in the 1240K dataset, restricting to transversions only to replicate key results (Supplementary Information). We started with a skeleton tree that fits the data with Denisova, Mbuti, Onge, Tianyuan and Loschbour and one admixture event. We then grafted on Mongolia_East_N, Jomon, Taiwan_Hanben, Chokhopani, and Boisman in turn, adding them consecutively to all possible edges in the tree and retaining only graph solutions that provided no differences of |Z|>3 between fitted and estimated statistics. We used the MSMC relative population split time to constrain models (the maximum discrepancy for this model is |Z|=2.8). Drifts along edges are multiplied by 1000. Dashed lines represent admixture. Deep population splits are not well constrained due to a lack of data from Upper Paleolithic East Asians.
445 To obtain insight into the formation of present-day Japanese archipelago populations,
446 we searched for groups that contribute most strongly to present-day Japanese through
447 admixture f3-statistics. The most strongly negative signals come from mixtures of Han
448 Chinese and ancient Jomon (f3(Japanese; Han Chinese, Jomon)) (Table S16). We can
449 model present-day Japanese as two-way mixtures of 84.3% Han Chinese and 15.7%
450 Jomon or 87.6% Korean and 12.4% Jomon (we cannot distinguish statistically
451 between these two sources; Table S17 and Table S18). This analysis by no means
452 suggests that the mainland ancestry in Japan was contributed directly by the Han
453 Chinese or Koreans themselves, but does suggest that it is from an ancestral
454 population related to those that contributed in large proportion to Han Chinese as well
455 as to Koreans for which we do not yet have ancient DNA data.
456
457 We used qpGraph35 to explore models with population splits and gene flow, and
458 tested their fit to the data by computing f2-, f3- and f4- statistics measuring allele
459 sharing among pairs, triples, and quadruples of populations, evaluating fit based on
460 the maximum |Z|-score comparing predicted and observed values. We further
461 constrained the models by using estimates of the relative population split times
462 between the selected pairs of populations based on the output of the MSMC
463 software50. While admixture graph modeling based on allele frequency correlation
464 statistics is not able to reject a model in which ancient Taiwan individuals and
465 Boisman share substantial ancestry with each other more recently than either does
466 with the ancestors of Chokopani and Core Tibetans, this model cannot be correct
467 because our MSMC analysis reveals that Core Tibetans (closely related to Chokopani)
468 and Ulchi (closely related to Boisman) share ancestry more recently in time on
469 average than either does with Ami (related to Taiwan_Hanben). This MSMC-based
470 constraint allowed us to identify a parsimonious working model for the deep history
471 of key lineages discussed in this study (Supplementary Information section 3:
472 qpGraph Modeling). Our fitted model (Figure 5), suggests that much of East Asian
473 ancestry today can be modelled as derived from two ancient populations: one from the
474 same lineage as the approximately ~40,000-year-old Tianyuan individual and the
475 other more closely related to Onge, with groups today having variable proportions of
476 ancestry from these two deep sources. In this model, the Mongolia_East_N and Amur
477 River Basin Boisman related lineages derive the largest proportion of their ancestry
478 from the Tianyuan-related lineage and the least proportion of ancestry from the Onge
479 related lineage compared with other East Asians. A sister lineage of
480 Mongolia_East_N is consistent with expanding into the Tibetan Plateau and mixing
481 with the local hunter-gatherers who represent an Onge-related branch in the tree. The
482 Taiwan Hanben are well modelled as deriving about 14% of their ancestry from a
483 lineage remotely related to Onge and the rest of their ancestry from a lineage that also
484 contributed to Jomon and Boisman on the Tianyuan side, a scenario that would
485 explain the observed affinity among Jomon, Boisman and Taiwan Hanben. We
486 estimate that Jomon individuals derived 45% of their ancestry from a deep basal
487 lineage on the Onge side. These results are consistent with the scenario a Late
488 Pleistocene coastal route of human migration linking Southeast Asia, the Japanese
489 Archipelago and the Russian Far East51. Due to the paucity of ancient genomic data
490 from Upper Paleolithic East Asians, there are limited constraints at present for
491 reconstructing the deep branching patterns of East Asian ancestral populations, and it
492 is certain that this admixture graph is an oversimplification and that additional
493 features of deep population relationships will be revealed through future work.
495 At the end of the last Ice Age, there were multiple highly differentiated populations in
496 East as well as West Eurasia, and it is now clear that these groups mixed in both
497 regions, instead of one population displacing the others. In West Eurasia, there were
498 at least four divergent populations each as genetically differentiated from each other
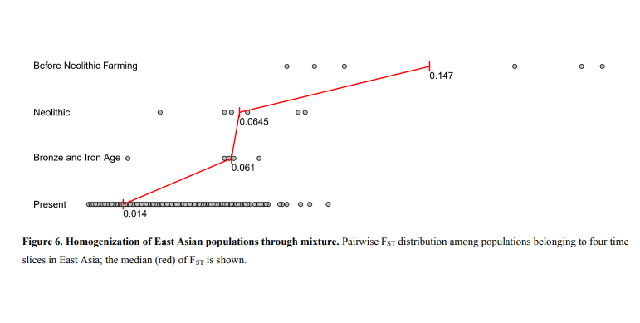
499 as Europeans and East Asians today (average FST=0.10), which mixed in the
500 Neolithic, reducing heterogeneity (average FST=0.03) and mixed further in the Bronze
501 Age and Iron Age to produce the present-relatively low differentiation that
502 characterizes modern West Eurasia (average FST=0.01)52. In East Eurasia, our study
503 suggests an analogous process, with the differentiation characteristic of the Amur
504 River Basin groups, Neolithic Yellow River farmers, and people related to those of
505 the Taiwan Iron Age (average FST=0.06 in our data) collapsing through mixture to
506 today’s relatively low differentiation (average FST=0.01-0.02) (Figure 6). A priority
507 should be to obtain ancient DNA data for the hypothesized Yangtze River population
508 (the putative source for the ancestry prevalent in the Southeast Asian Cluster of
509 present-day groups), which should, in turn, make it possible to test and further extend
510 these models, and in particular to understand if dispersals of people in Southeast Asia
511 do or do not correlate to ancient movements of people.
 |
|
|
|
Post by Admin on Apr 2, 2020 5:50:36 GMT
A dynamic 6,000-year genetic history of Eurasia’s Eastern Steppe Choongwon Jeong1,2,†,*, Ke Wang1,†, Shevan Wilkin3, William Timothy Treal Taylor3,4
61 Summary
62
63 The Eastern Eurasian Steppe was home to historic empires of nomadic pastoralists, including
64 the Xiongnu and the Mongols. However, little is known about the region’s population history.
65 Here we reveal its dynamic genetic history by analyzing new genome-wide data for 214
66 ancient individuals spanning 6,000 years. We identify a pastoralist expansion into Mongolia
67 ca. 3000 BCE, and by the Late Bronze Age, Mongolian populations were biogeographically
68 structured into three distinct groups, all practicing dairy pastoralism regardless of ancestry.
69 The Xiongnu emerged from the mixing of these populations and those from surrounding
70 regions. By comparison, the Mongols exhibit much higher Eastern Eurasian ancestry,
71 resembling present-day Mongolic-speaking populations. Our results illuminate the complex
72 interplay between genetic, sociopolitical, and cultural changes on the Eastern Steppe.
73
74 Keywords
75
76 Human population history, ancient DNA, migration, nomadic pastoralists, Eastern Steppe,
77 Mongolia, Xiongnu Empire, Mongol Empire
79 Introduction
80
81 Recent paleogenomic studies have revealed a dynamic population history on the Eurasian
82 Steppe, with continental-scale migration events on the Western Steppe coinciding with Bronze
83 Age transformations of Europe, the Near East, and the Caucasus (Allentoft et al., 2015; de
84 Barros Damgaard et al., 2018; Damgaard et al., 2018; Haak et al., 2015; Mathieson et al., 2015;
85 Wang et al., 2019). However, despite advances in understanding the genetic prehistory of the
86 Western Steppe, the prehistoric population dynamics on the Eastern Steppe remain poorly
87 understood (de Barros Damgaard et al., 2018; Jeong et al., 2018). The Eastern Steppe is a great
88 expanse of grasslands, forest-steppe, and desert-steppe extending more than 2,500 km from the
89 Altai-Sayan mountain range in the west to northeastern China in the east (Fig. 1). While also
90 covering parts of modern-day China and Russia, most of the Eastern Steppe falls within the
91 national boundaries of present-day Mongolia (Fig. S1). The Eastern Steppe has been occupied
92 since the early Upper Paleolithic (ca. 34,000 cal BP) (Devièse et al., 2019), and recent
93 paleogenomic studies suggest that the eastern Eurasian forest-steppe zone was genetically
94 structured during the Pre-Bronze and Early Bronze Age periods, with a strong west-east
95 admixture cline of ancestry stretching from Botai in central Kazakhstan to Lake Baikal in
96 southern Siberia to Devil’s Gate Cave in the Russian Far East (de Barros Damgaard et al.,
97 2018; Jeong et al., 2018; Sikora et al., 2019; Siska et al., 2017).
98
99 During the Bronze Age, the multi-phased introduction of pastoralism drastically changed
100 lifeways and subsistence on the Eastern Steppe (Honeychurch, 2015; Kindstedt and Ser-Od,
101 2019). A recent large-scale paleoproteomic study has confirmed milk consumption in
102 Mongolia prior to 2500 BC by individuals affiliated with the Afanasievo (3000 BCE) and
103 Chemurchek (2750-1900 BCE) cultures (Wilkin et al., 2019). Although Afanasievo groups in
104 the Upper Yenisei region have been genetically linked to the Yamnaya culture of the Pontic
105 Caspian steppe (ca. 3300-2200 BCE) (Allentoft et al., 2015; Morgunova and Khokhlova,
106 2013), the origins of the Chemurchek have been controversial (Kovalev, 2014). Once
107 introduced, ruminant dairying became widespread by the Middle/Late Bronze Age (MLBA,
108 here defined as 1900-900 BCE), being practiced in the west and north at sites associated with
109 Deer Stone-Khirigsuur Complex (DSKC) and in the east in association with the Ulaanzuukh
110 culture (Jeong et al., 2018; Wilkin et al., 2019). The relationships between DSKC and
111 Ulaanzuukh groups are poorly understood, and little is known about other MLBA burial
112 traditions in Mongolia, such as the Mönkhkhairkhan and Baitag (See supplementary materials).
113 By the end of the second millennium BCE, the previously powerful MLBA cultures were in
114 decline, and political power shifted during the Early Iron Age to the Slab Grave culture (ca.
115 1000-300 BCE), whose burials often incorporate uprooted materials from DSKC monuments
116 (Honeychurch, 2015), and to the Uyuk culture (ca. 700-200 BCE) of the Sayan mountains to
117 the northwest (also known as the Aldy-Bel culture), who had strong cultural ties to the Pazyryk
118 (ca. 500-200 BCE) and Saka (ca. 900-200 BCE) cultures of the Altai and eastern Kazakhstan
119 (Savinov, 2002; Tseveendorj, 2007).
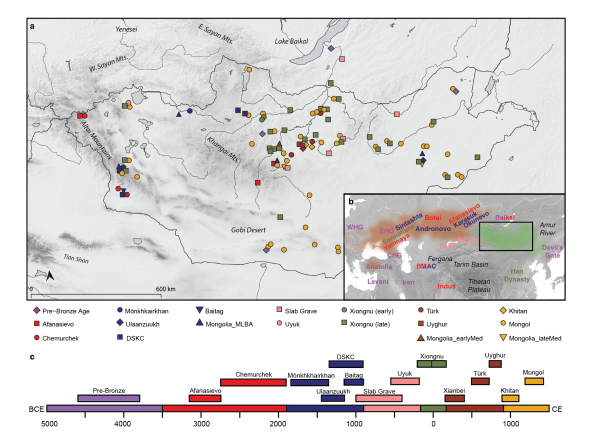 Fig. 1. Overview of ancient populations and time periods
121 From the late first millennium BC onwards, a series of hierarchical and centrally organized
122 empires arose on the Eastern Steppe, notably the Xiongnu (209 BCE-98 CE), Türkic (552-742
123 CE), Uyghur (744-840 CE), and Khitan (916-1125 CE) empires. The Mongol empire, emerging
in the 13th 124 century CE, was the last and most expansive of these regimes, eventually controlling
125 vast territories and trade routes stretching from China to the Mediterranean. However, due to a
126 lack of large-scale genetic studies, the origins and relationships of the people who formed these
127 states, including both the ruling elites and local commoners, remain obscure (See
128 supplementary materials).
130 To clarify the population dynamics on the Eastern Steppe since prehistory, we generated and
131 analyzed genome-wide genetic datasets for 214 individuals from 85 Mongolian and 3 Russian
132 sites spanning approximately 6,000 years of time (ca. 4600 BCE to 1400 CE) (Tables S1-S9
133 and Fig. S2-S8) (See supplementary materials). To this, we added recently published genomic
134 data for 18 Bronze Age individuals from northern Mongolia (Jeong et al., 2018), as well as
135 datasets from neighboring ancient populations in Russia and Kazakhstan (de Barros Damgaard
136 et al., 2018; Damgaard et al., 2018; Narasimhan et al., 2019; Sikora et al., 2019; Unterländer
137 et al., 2017) (Table S10), which we analyze together with worldwide modern reference
138 populations (Table S11; Fig. S4). We also generated 24 new accelerator mass spectrometry
139 dates (Table S12), supplementing 81 previously published radiocarbon dates from Mongolia
140 (Jeong et al., 2018; Taylor et al., 2019), for a total of 105 directly dated individuals in this study
141 (See supplementary materials).
|
|
|
|
Post by Admin on Apr 3, 2020 6:42:51 GMT
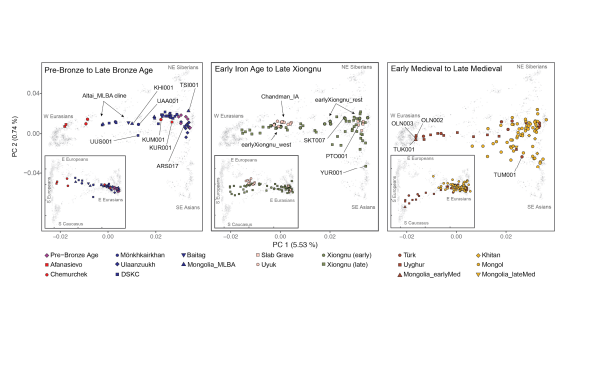 Fig. 2. Genetic structure of Mongolia through time
143 Results
144
145 Pre-Bronze Age population structure and the arrival of pastoralism
146
147 Throughout the mid-Holocene and before the Bronze Age, the Eastern Steppe was sparsely
148 populated by hunter-gatherers, whose lives and activities are recorded in petroglyphs
149 throughout the region (Jacobson-Tepfer and Meacham, 2010; Kindstedt and Ser-Od, 2019). In
150 this study, we analyzed six pre-Bronze Age individuals from three sites dating to the fifth and
151 fourth millennia BCE: one from eastern Mongolia (SOU001, “eastMongolia_preBA”, 4686-
152 4495 cal. BCE), one from central Mongolia (ERM003, “centralMongolia_preBA”, 3781-3643
153 cal. BCE), and four from the eastern Baikal region (“Fofonovo_EN”). By comparing these
154 genomes to previously published ancient and modern data across Eurasia (Fig. 2) (see Methods
155 and Materials), we found that they are most closely related to contemporaneous hunter
156 gatherers from the western Baikal region (“Baikal_EN”, 5200-4200 BCE) and the Russian Far
157 East (“DevilsCave_N”, ca. 5700 BCE), filling in the geographic gap in the distribution of this
158 genetic profile (Fig. 3a). We refer to this profile as “Ancient Northeast Asian” (ANA) to reflect
159 its geographic distribution relative to another widespread mid-Holocene genetic profile known
160 as “Ancient North Eurasian” (ANE), which is found among the Pleistocene hunter-gatherers
161 of the Mal’ta (ca. 24500-24100 BP) and Afontova Gora (ca. 16900-16500 BP) sites in Siberia
162 (Fu et al., 2016; Raghavan et al., 2015) and the horse-herders of Botai, Kazakhstan (ca. 3500-
163 3300 BCE) (de Barros Damgaard et al., 2018). In PCA (Fig. 2), ancient ANA individuals fall
164 close to the cluster of present-day Tungusic- and Nivkh-speaking populations from the lower
165 Amur River region in northeast Asia, indicating that their genetic profile is still present in
166 indigenous populations of the Far East today (Fig. S5). EastMongolia_preBA is genetically
167 indistinguishable from DevilsCave_N (Fig. 3a; Table S13; Fig. S9-S10), whereas
168 Fofonovo_EN and the slightly later centralMongolia_preBA both derive a minority (12-17%)
169 of their ancestry from ANE-related (Botai-like) groups with the remainder of their ancestry
170 (83-87%) characterized as ANA (Fig. 3a; Table S13). Reanalyzing published data from the
171 western Baikal early Neolithic Kitoi culture (Baikal_EN) and the early Bronze Glazkovo
172 culture (Baikal_EBA) (de Barros Damgaard et al., 2018), we find that they have similar
173 ancestry profiles and a slight increase in ANE ancestry through time (from 6.4% to 20.1%)
174 (Fig. 3a). Overall, these data suggest broader genetic shifts in hunter-gatherer ancestry at this
175 time.
176
177 Pastoralism in Mongolia is often assumed to have been introduced by the eastward expansion
178 of Western Steppe cultures (e.g., Afanasievo) via either the Upper Yenisei and Sayan mountain
179 region to the northwest of Mongolia or through the Altai mountains in the west (Janz et al.,
180 2017). Although the vast majority of Afanasievo burials found to date are located in the Altai
181 mountains and Upper Yenisei regions outside of Mongolia (Honeychurch, 2017), an Early
182 Bronze Age (EBA) site in the southern Khangai Mountains of central Mongolia has yielded
183 Afanasievo-style graves with proteomic evidence of ruminant milk consumption (Wilkin et al.,
184 2019). Analyzing two of these individuals (Afanasievo_Mongolia, 3112-2917 cal. BCE), we
185 find that their genetic profiles are indistinguishable from that of published Afanasievo
186 individuals from the Yenisei region (Allentoft et al., 2015; Narasimhan et al., 2019) (Fig. 2,
187 Fig. S11), and thus these two Afanasievo individuals confirm that the EBA expansion of
188 Western Steppe herders (WSH) did not stop at the mountain ranges separating the Western and
189 Eastern Steppes (Allentoft et al., 2015), but rather extended a further 1500 km eastwards into
190 the heart of central Mongolia (Fig. 3a).
192 Although the Afanasievo-related expansion may have introduced dairy pastoralism into
193 Mongolia (Janz et al., 2017), the people themselves do not seem to have left a long-lasting
194 genetic footprint. An Afanasievo-style burial (Afanasievo_KUR001) excavated in the
195 Mongolian Altai and dating to approximately 500 years later (2618-2487 cal. BCE), was found
196 to have a distinct (Fig. 2) and mostly ANA (~90%) derived ancestry (Fig. 3a), with only a
197 minor component tracing back to the Afanasievo (~10%; Table S14; Fig. S11).
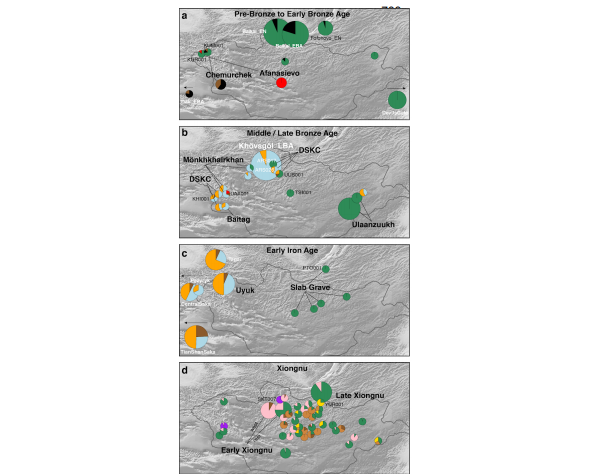 Fig. 3. Genetic changes in the Eastern Steppe across time characterized by 736 qpAdm.
198
199 The succeeding EBA Chemurchek culture (2750-1900 BCE), a ruminant dairying society
200 (Wilkin et al., 2019) whose mortuary features include stone slabs and anthropomorphic stelae,
201 has also been purportedly linked to WSH migrations (Kovalev and Erdenebaatar, 2009).
202 Chemurchek graves are found throughout the Altai and in the Dzungar Basin in Xinjiang, China
203 (Jia and Betts, 2010). We analyzed two Chemurchek individuals from the southern Altai site
204 of Yagshiin Huduu and one from Khundii Gobi (KUM001) in the northern Altai. Compared to
205 Afanasievo_Mongolia, the Yagshiin Huduu individuals also show a high degree of Western
206 ancestry but are displaced in PCA (Fig. 2), having also a strong genetic affinity with ANE207 related ancient individuals such as AfontovaGora3 (AG3), West_Siberia_N, and Botai (Fig.
208 3a; Fig. S9, S11). We find that these Chemurchek individuals (Chemurchek_Altai) are
209 genetically similar to Dali_EBA (Fig. 3a), a contemporaneous individual from eastern
210 Kazakhstan (Narasimhan et al., 2019). The genetic profiles of both the Yagshiin Huduu and
211 Dali_EBA individuals are well fitted by two-way admixture models with Botai (60-78%) and
212 groups with ancient Iranian-related ancestry, such as Gonur1_BA from Gonur Tepe, a key EBA
213 site of the Bactria-Margiana Archaeological Complex (BMAC) (22-40%; Table S15; Fig. 3a),
214 and ancient individuals from post-BMAC Bronze Age sites in southeastern Uzbekistan (Table
215 S15). Although minor genetic contributions from the Afanasievo-related groups cannot be
216 excluded, Iranian-related ancestry is required for all fitting models, and this admixture is
217 estimated to have occurred 12±6 generations earlier (~336±168 years; Fig. S12) when modeled
218 using DATES (Narasimhan et al., 2019). Because all proxy source populations used in this
219 modeling are quite distant in either time or space from the EBA Altai, the proximate
220 populations contributing to the Chemurchek cannot be precisely identified, but future research
221 in genetically unsampled regions such as Xinjiang may provide important clues.
222
223 In the northern Altai, the third Chemurchek individual in our study (KUM001) has additional
224 northeast Asian ancestry, similar to levels seen in Afanasievo_KUR001 (Fig. 3a). KUM001
225 can be modeled as a mixture of Chemurchek-related (~30%) and ANA ancestry (~70%) (Table
226 S15). The high proportions of local ANA ancestry in KUR001 and KUM001 suggest that the
227 arrival of Afanasievo and later Chemurchek pastoralists did not necessarily displace local Altai
228 peoples, despite the strong cultural and economic impact of these herders. Indeed, the cultural
229 features that they first introduced, such as mortuary mound building and dairy pastoralism,
230 continue to the present day. However, unlike in Europe, where migrating EBA steppe herders
231 had a transformative and lasting genetic impact on local populations (Allentoft et al., 2015;
232 Haak et al., 2015; Mathieson et al., 2018), we find that neither Afanasievo nor Chemurchek
233 groups left significant or enduring genetic traces into the subsequent Middle and Late Bronze
234 Ages (MLBA).
236 Bronze Age emergence of a tripartite genetic structure
237
238 Previously, we reported a shared genetic profile among EBA western Baikal hunter-gatherers
239 (Baikal_EBA) and Late Bronze Age (LBA) pastoralists in northern Mongolia
240 (Khövsgöl_LBA) (Jeong et al., 2018). This genetic profile, composed of major and minor ANA
241 and ANE ancestry components, respectively, is also shared with the earlier eastern Baikal
242 (Fofonovo_EN) and Mongolian (centralMongolia_preBA) groups analyzed in this study (Fig.
243 3a,b), suggesting a regional persistence of this genetic profile for nearly three millennia.
244 Centered in northern Mongolia, this genetic profile is distinct from that of other Bronze Age
245 groups. Overall, we find three distinct and geographically structured gene pools in LBA
246 Mongolia, with the Khövsgöl_LBA population representing one of them (Fig. 3b). The other
247 two, which we refer to as “Altai_MLBA” and “Ulaanzuuk_SlabGrave”, are described below.
248
249 During the MLBA (1900-900 BCE), as grasslands expanded in response to climate change,
250 new pastoralist cultures expanded out of inner-montane regions and across the Eastern Steppe
251 (Kindstedt and Ser-Od, 2019). This period is also notable for the first regional evidence horse
252 milking (ca. 1200 BCE; (Wilkin et al., 2019)), and a dramatic intensification of horse use,
253 including the emergence of mounted horseback riding that would have substantially extended
254 the accessibility of remote regions of the steppe. Today, horse milking is exclusively associated
255 with alcohol production (Bat-Oyun et al., 2015), and this first appearance of horse milk may
256 mark the origin of this social tradition on the Eastern Steppe. In the Altai-Sayan region, dairy
257 pastoralists associated with the Mönkhkhairkhan, Deer Stone-Khirigsuur Complex (DSKC),
258 and Baitag cultures (Altai_MLBA, n=7), all show clear genetic evidence of admixture between
259 a Khövsgöl_LBA-related ancestry and a Sintashta-related WSH ancestry (Fig. 3b). Overall,
260 they form an “Altai_MLBA” cline on PCA between Western Steppe groups and the
261 Baikal_EBA/Khövsgöl_LBA cluster (Fig. 2), with their position varying on PC1 according to
262 their level of Western ancestry (Table S16).
263
264 This is the first appearance on the Eastern Steppe of a Sintashta-like ancestry (frequently
265 referred to as “steppe_MLBA” in previous studies), which is distinct from prior Western
266 ancestries present in the Afanasievo and Chemurchek populations, and which instead shows a
267 close affinity to European Corded-Ware populations and later Andronovo-associated groups,
268 such as the Sintashta (Allentoft et al., 2015). In Khovd province, individuals belonging to
269 Mönkhkhairkhan and DSKC cultures (SBG001 and BER002, respectively) have a similar
270 genetic profile that is best modeled as an equal mixture of Khövsgöl_LBA and Sintashta (Fig.
271 3b; Table S16). This genetic profile matches that previously described for a genetic outlier in
272 northern Mongolia that deviated from the Khövsgöl_LBA cluster in a previous study (ARS026;
273 (Jeong et al., 2018)). An additional four Altai_MLBA individuals belonging to Baitag
274 (ULI003), DSKC (ULI001), and an unclassified culture (BIL001, ULZ001) also fit this
275 admixture model with varying admixture proportions (Table S16). Taken together, the
276 Altai_MLBA cline reveals the ongoing mixture of two source populations: a
277 Sintashta/Andronovo-related WSH population and a local population represented by
278 Khövsgöl_LBA. The admixture is estimated to have occurred only 9±2 generations (~250
279 years) before the individuals analyzed in this study, a finding consistent with their
280 heterogeneous ancestry proportions (Fig. S12 and S13). Because the Sintashta culture (ca.
281 2800-1600 BCE) is associated with novel transportation technologies, such as horse-drawn
282 chariots (Anthony, 2010), the appearance of this ancestry profile on the Eastern Steppe suggests
283 that heightened mobility capabilities played an important role in linking diverse populations
284 across the Eurasian Steppe.
285
286 Three MLBA individuals in our dataset present genetic profiles that cannot be fully explained
287 by the Altai_MLBA cline. The Altai Mönkhkhairkhan individual UAA001 matches with 3-
288 way admixture models using Khövsgöl_LBA/Baikal_EBA as the eastern Eurasian source,
289 Afanasievo as the WSH source (but not Sintashta), and adding Gonur1_BA as a third source
290 (Table S16). Thus, a minor Iranian-related ancestry component is necessary to model this
291 individual’s genetic profile, as reflected by his displacement along PC2 in Fig. 2. We also find
292 that the culturally unclassified Altai individual KHI001 may need additional ancestry from
293 Gonur1_BA to improve the existing 2-way admixture model. Last, UUS001, despite coming
294 from a DSKC context in Khövsgöl province, fits better with a 3-way model of using
295 eastMongolia_preBA and Gonur1_BA with Sintashta as the WSH source. Although cultural
296 differences may have existed among the major MLBA mortuary traditions of the Altai and
297 northern Mongolia (Mönkhkhairkhan, DSKC, and Baitag), they do not form distinct genetic
298 groups. Rather, these mortuary traditions encompass individuals of disparate and recently
299 admixed ancestry, and the proliferation and variability of mortuary features at this time likely
300 reflects this demographic dynamism.
302 The populations making up the heterogeneous Altai_MLBA cline left descendants in the Altai303 Sayan region, who we later identify at the Uyuk site of Chandman Mountain (“Chandman_IA”,
304 ca. 400-200 BCE) in northwestern Mongolia during the Early Iron Age (EIA). Nine
305 Chandman_IA individuals form a tight cluster on PCA at the end of the previous Altai_MLBA
306 cline away from Khövsgöl_LBA cluster (Fig. 2). During the EIA, the Uyuk (also known as the
307 Aldy-Bel or Sagly-Bazhy culture) were pastoralists largely centered in the Upper Yenisei
308 region of present-day Tuva, and together with the Pazyryk, with whom they share log chamber
309 tombs and other features, they formed part of a broader Saka cultural phenomenon that
310 stretched across the Western Steppe, the Tarim Basin, and the Upper Yenisei (Parzinger, 2006).
311 In addition to dairying, many Saka groups, including EIA individuals at the Chandman
312 Mountain site, also cultivated millet (Murphy et al., 2013; Ventresca Miller and Makarewicz,
313 2019; Wilkin et al.), a practice that had been earlier adopted by many southern steppe and
314 Central Asian populations during the westward spread of millet from China to the Caucasus
315 during the second millennium BCE (Ventresca Miller and Makarewicz, 2019).
316
317 We find that EIA Saka populations systematically deviate from the earlier Altai_MLBA cline,
318 requiring a third ancestral component (Fig. 3c). The appearance of this ancestry, related to
319 populations of Central Asia (Caucasus/Iranian Plateau/Transoxiana regions) including BMAC
320 (Narasimhan et al., 2019), is clearly detected in the Iron Age groups such as Central Saka
321 (Damgaard et al., 2018), TianShan Saka (Damgaard et al., 2018), Tagar (Damgaard et al.,
322 2018), and Chandman_IA (Tables S17-S18), while absent in the earlier DSKC and Karasuk
323 groups (Tables S16-S17). This third component makes up 6-24% of the ancestry in these Iron
324 Age groups, and the date of admixture in Chandman_IA is estimated ~17±4 generations earlier,
325 ca. 750 BCE, which postdates the collapse of the BMAC ca. 1600 BCE and slightly predates
326 the formation of the Persian Achaemenid empire ca. 550 BCE (Fig. S12, S13). We suggest that
327 this Iranian-related genetic influx was mediated by increased contact and mixture with
328 agropastoralist populations in the region of Transoxiana (Turan) and Fergana during the LBA
329 to EIA transition. The widespread emergence of horseback riding during the late second and
330 early first millennium BCE (Drews, 2004), and the increasing sophistication of horse transport
331 thereafter, likely contributed to increased population contact and the dissemination of this
332 Iranian-related ancestry onto the steppe. Our results do not exclude additional spheres of
333 contact, such as increased mobility along Inner Asian Mountain Corridor, which could have
334 also introduced this ancestry into the Altai via Xinjiang starting in the Bronze Age (Frachetti,
335 2012).
336
337 In contrast to the MLBA and EIA cultures of the Altai and northern Mongolia, different burial
338 traditions are found in the eastern and southern regions of Mongolia (Honeychurch, 2015),
339 notably the LBA Ulaanzuukh (1450-1150 BCE) and EIA Slab Grave (1000-300 BCE) cultures.
340 In contrast to other contemporaneous Eastern Steppe populations, we find that individuals
341 associated with these burial types show a clear northeastern-Eurasian (ANA-related) genetic
342 profile lacking both ANE and WSH admixture (Fig. 2; Fig. 3c; Fig. S7). Both groups were
343 ruminant pastoralists, and the EIA Slab Grave culture also milked horses (Wilkin et al., 2019).
344 The genetic profiles of Ulaanzuukh and Slab Grave individuals are genetically
345 indistinguishable (Fig. 2 and Table S16), consistent with the archaeological hypothesis that the
346 Slab Grave tradition emerged out of the LBA Ulaanzuukh (Honeychurch, 2015; Khatanbaatar,
347 2019). Both groups are also indistinguishable from the earlier eastMongolia_preBA individual
348 dating to ca. 4600 BCE, suggesting a long-term (>4,000 year) stability of this prehistoric
349 eastern Mongolian gene pool (Table S16). In subsequent analyses, we merged Ulaanzuukh and
350 Slab Grave into a single genetic group (“Ulaanzuukh_SlabGrave”). The
351 Ulaanzuukh_SlabGrave genetic cluster is the likely source of the previously described DSKC
352 eastern outlier from Khövsgöl province (ARS017) (Jeong et al., 2018), as well as a culturally
353 unclassified individual (TSI001) from central Mongolia who dates to the LBA-EIA transition
354 (Figs. 2 and 3c; Table S16). In addition, the Mönkhkhairkhan individual KHU001 from
355 northwest Mongolia has a non-negligible amount of Ulaanzuukh_SlabGrave ancestry in
356 addition to his otherwise Baikal_EBA ancestry (Fig. 3c; Table S16). While these three
357 individuals attest to occasional long-distance contacts between northwestern and eastern
358 Mongolia during the LBA, we find no evidence of Ulaanzuukh_SlabGrave ancestry in the
359 Altai, and the overall frequency of the Ulaanzuukh_SlabGrave genetic profile outside of
360 eastern and southern Mongolia during the MLBA is very low. During the EIA, the Slab Grave
361 culture expanded northwards, often disrupting and uprooting former DSKC graves in their path
362 (Honeychurch, 2015), and it ultimately reached as far north as the eastern Baikal region, which
363 is reflected in the genetic profile of the Slab Grave individual PTO001 in this study (Fig. 3c).
364 Overall, our findings reveal a strong east-west genetic division among Bronze Age Eastern
365 Steppe populations through the end of the Early Iron Age.
|
|
|
|
Post by Admin on Apr 4, 2020 5:41:34 GMT
367 The Xiongnu Empire, the rise of the first imperial steppe polity
368
369 Arising from the prehistoric populations of the Eastern Steppe, large-scale polities began to
370 develop during the late first millennium BCE. The Xiongnu was the first historically
371 documented empire founded by pastoralists, and its establishment is considered a watershed
372 event in the sociopolitical history of the Eastern Steppe (Brosseder and Miller, 2011;
373 Honeychurch, 2015). The Xiongnu held political dominance in East and Central Asia from the
374 third century BCE through the first century CE. The cultural, linguistic and genetic make-up
375 of the people who constituted the Xiongnu empire has been of great academic interest, as has
376 their relationship to other contemporaneous and subsequent nomadic groups on the Eastern
377 Steppe. Here we report genome-wide data for 60 Xiongnu-era individuals from across
378 Mongolia and dating from 350 BCE to 130 CE, thus spanning the entire period of the Xiongnu
379 empire. Although most individuals date to the late Xiongnu period (after 50 BCE), 13
380 individuals predate 100 BCE and include 12 individuals from the northern early Xiongnu
381 frontier sites of Salkhityn Am (SKT) and Atsyn Gol (AST) and one individual from the early
382 Xiongnu site of Jargalantyn Am (JAG) in eastern Mongolia.
383
384 We observe two distinct demographic processes that contributed to the formation of the early
385 Xiongnu. First, half of the individuals (n=6) form a genetic cluster (earlyXiongnu_west)
386 resembling that of Chandman_IA of the preceding Uyuk culture from the Altai-Sayan region
387 (Fig. 2). They derive 91.8±1.9% of their ancestry from Chandman_IA with the remainder
388 attributed to additional Iranian-related ancestry, which we model using BMAC as a proxy (Fig.
389 3d; Table S19). This suggests that the low-level Iranian-related gene flow identified among the
390 Chandman Mountain Uyuk during the EIA likely continued during the second half of the first
391 millennium BCE, spreading across western and northern Mongolia. Second, six individuals
392 (“earlyXiongnu_rest”) fall intermediate between the earlyXiongnu_west and
393 Ulaazuukh_SlabGrave clusters; four carry varying degrees of earlyXiongnu_west (39-75%)
394 and Ulaazuukh_SlabGrave (25-61%) related ancestry, and two (SKT004, JAG001) are
395 indistinguishable from the Ulaanzuuk_SlabGrave cluster (Fig. 3d; Tables S19-S20). This
396 genetic cline linking the earlyXiongnu_west and Ulaanzuukh_SlabGrave gene pools signifies
397 the unification of two deeply diverged and distinct lineages on the Eastern Steppe - between
398 the descendants of the DSKC, Mönkhkhairkhan, Baitag, and Uyuk cultures in the west and the
399 descendants of the Ulaanzuuk and Slab Grave cultures in the east - and even tracing back before
400 the Bronze Age, when genetic differentiation is seen even among the northern/western and
401 eastern hunter-gatherers. Overall, the low-level influx of Iranian-related gene flow continuing
402 from the previous Uyuk culture and the sudden appearance of a novel east-west mixture uniting
403 the gene pools of the Eastern Steppe are the two defining demographic processes associated
404 with the rise of the Xiongnu.
405
406 Among late Xiongnu individuals, we find even higher genetic heterogeneity (Fig. 2), and their
407 distribution on PC indicates that the two demographic processes evident among the early
408 Xiongnu continued into the late Xiongnu period, but with the addition of new waves and
409 complex directions of gene flow. Of the 47 late Xiongnu individuals, half (n=26) can be
410 adequately modelled by the same admixture processes seen among the early Xiongnu: 22 as a
411 mixture of Chandman_IA+Ulaanzuuk_SlabGrave, two (NAI002, TUK002) as a mixture of
412 either Chandman_IA+BMAC or Chandman_IA+Ulaanzuuk_SlabGrave+BMAC, and two
413 (TUK003, TAK001) as a mixture of either earlyXiongnu_west+Ulaanzuukh_SlabGrave or
414 earlyXiongnu_west+Khovsgol_LBA (Fig 3d). A further two individuals (TEV002, BUR001)
415 also likely derive their ancestry from early Xiongnu gene pool, although the p-value of their
416 models is slightly lower than the 0.05 threshold (Table S20). However, a further 11 late
417 Xiongnu with the highest proportions of western Eurasian affinity along PC1 cannot be
418 modelled using BMAC or any other ancient Iranian-related population. Instead, they fall on a
419 cluster of ancient Sarmatians from various locations in the western and central steppe (Fig. 2).
420
421 Admixture modeling confirms the presence of a Sarmatian-related gene pool among the late
422 Xiongnu: three individuals (UGU010, TMI001, BUR003) are indistinguishable from
423 Sarmatian, two individuals (DUU001, BUR002) are admixed between Sarmatian and BMAC,
424 three individuals (UGU005, UGU006, BRL002) are admixed between Sarmatian and
425 Ulaanzuuk_SlabGrave, and three individuals (NAI001, BUR004, HUD001) require Sarmatian,
426 BMAC and Ulaanzuuk_SlabGrave (Fig. 3d; Table S20. In addition, eight individuals with the
427 highest eastern Eurasian affinity along PC1 are distinct from both the Ulaanzuuk_SlabGrave
428 and Khövsgöl_LBA genetic profiles, showing affinity along PC2 towards present-day people
429 from East Asia further to the south (Fig. 2). Six of these individuals (EME002, ATS001,
430 BAM001, SON001, TUH001, YUR001) are adequately modeled as a mixture of
431 Ulaanzuuk_SlabGrave and Han (Table S19-S20), and YUR001 in particular exhibits a close
432 genetic similarity to two previously published Han empire soldiers (Damgaard et al., 2018),
433 whose genetic profile we refer to as “Han_2000BP” (Table S20). The remaining two
434 individuals (BRU001, TUH002) are similar but also require the addition of Sarmatian ancestry
435 (Fig. 3d, Table S20). The late Xiongnu are thus characterized by two additional demographic
436 processes that distinguish them from the early Xiongnu: gene flow from a new Sarmatian
437 related Western ancestry source and intensified interaction and mixture with people of the
438 contemporaneous Han empire of China. Together, these results match well with historical
439 records documenting the political influence that the Xiongnu exercised over their neighbors,
440 including the Silk Road kingdoms of Central Asia and Han Dynasty China, as well as purported
441 migrations both in and out of Mongolia (Miller, 2014). Overall, the Xiongnu period can be
442 characterized as one of expansive and extensive gene flow that began by uniting the gene pools
443 of western and eastern Mongolia and ended by uniting the gene pools of western and eastern
444 Asia.

446 Fluctuating genetic heterogeneity in the post-Xiongnu polities
447
448 After the collapse of the Xiongnu empire ca. 100 CE, a succession of nomadic pastoralist
449 regimes rose and fell over the next several centuries across the politically fragmented Eastern
450 Steppe: Xianbei (ca. 100-250 CE), Rouran (ca. 300-550 CE), Türkic (552-742 CE), and Uyghur
451 (744-840 CE). Although our sample representation for the Early Medieval period is uneven,
452 consisting of one unclassified individual dating to the Xianbei or Rouran period (TUK001), 8
453 individuals from Türkic mortuary contexts, and 13 individuals from Uyghur cemeteries, it is
454 clear that these individuals have genetic profiles that differ from the preceding Xiongnu period,
455 suggesting new sources of gene flow into Mongolia at this time that displace them along PC3
456 (Fig. 2). Individual TUK001 (250-383 cal. CE), whose burial was an intrusion into an earlier
457 Xiongnu cemetery, has the highest western Eurasian affinity. This ancestry is distinct from that
458 of the Sarmatians, and closer to ancient populations with BMAC/Iranian-related ancestry (Fig.
459 2). Among the individuals with the highest eastern Eurasian affinity, two Türkic- and one
460 Uyghur-period individual (ZAA004, ZAA002, OLN001.B) are indistinguishable from the
461 Ulaanzuuk_SlabGrave cluster. Another individual (TUM001), who was recovered from the
462 tomb ramp of an elite Türkic-era emissary of the Tang Dynasty, has a high proportion of Han463 (78.1±1.5%) (Fig. 3e) and especially Han_2000BP-related ancestry (84±1.5%) (Table S21).
464 This male, buried with two dogs, was likely a Chinese attendant sacrificed to guard the tomb
465 entrance (Ochir et al., 2013). The remaining 17 Türkic and Uyghur individuals show
466 intermediate genetic profiles (Fig. 3e).
467
468 The high genetic heterogeneity of the Early Medieval period is vividly exemplified by twelve
469 individuals from the Uyghur period cemetery of Olon Dov (OLN; Fig. 2), in the vicinity of the
470 Uyghur capital of Ordu-Baliq. Six of these individuals came from a single tomb (grave 19), of
471 whom only two are related (OLN002 and OLN003, second-degree; Table S7); the absence of
472 closer kinship ties raises questions about the function of such tombs and the social relationships
473 of those buried within them. Most Uyghur period individuals exhibit a high, but variable,
474 degree of west Eurasian ancestry - best modeled as a mixture of Alans (a historic nomadic
475 pastoral group likely descended from the Sarmatians and contemporaries of the Huns
476 (Bachrach, 1973)) and an Iranian-related (BMAC-related) ancestry - together with
477 Ulaanzuukh_SlabGrave (ANA-related) ancestry (Fig. 3e). The admixture dates estimated for
478 the ancient Türkic and Uyghur individuals in this study correspond to ca. 500 CE: 8±2
479 generations before the Türkic individuals and 12±2 generations before the Uyghur individuals
480 (represented by ZAA001 and Olon Dov individuals).
|
|
|
|
Post by Admin on Apr 4, 2020 21:09:14 GMT
482 Rise of the Mongol empire
483
484 After the fall of the Uyghur empire in the mid-ninth century, the Khitans of northeast China
485 established the powerful Liao Dynasty in 916 CE. Although few Khitan sites are known within
486 Mongolia, the Khitans controlled large areas of the Eastern Steppe and are recorded to have
487 relocated people within their conquered territories (Kradin and Ivliev, 2008). Our study
488 includes three Khitan individuals (ZAA003, ZAA005, ULA001) from Bulgan province, all of
489 whom have a strongly eastern Eurasian (ANA and Han_2000BP-related) genetic profile (Fig.
490 2), with <10% west Eurasian ancestry (Fig. 3f; Table S22). This may reflect the northeastern
491 Asian origin of the Mongolic-speaking Khitan, but a larger sample size is required to
492 adequately characterize the genetic profile of Khitan populations within Mongolia. In 1125 CE,
493 the Khitan empire fell to the Jurchen’s Jin Dynasty, which was then conquered in turn by the
494 Mongols in 1234 CE.
495
496 At its greatest extent, the Mongol empire (1206-1368 CE) spanned nearly two thirds of the
497 Eurasian continent. It was the world’s largest contiguous land empire, and the cosmopolitan
498 entity comprised diverse populations that flowed into the steppe heartland. We analyzed 62
499 Mongol-era individuals whose burials are consistent with those of low-level, local elites. No
500 royal or regional elite burials were included, nor were individuals from the cosmopolitan
501 capital of Karakorum. Although we find that Mongol-era individuals were diverse, they exhibit
502 a much lower genetic heterogeneity compared to Xiongnu-era individuals (Fig. 2), and they
503 almost entirely lack the residual ANE-related ancestry (in the form of Chandman_IA and
504 Khövsgöl_LBA) that had been present among the Xiongnu and earlier northern/western
505 MLBA cultures. On average, Mongol period individuals have a much higher eastern Eurasian
506 affinity than previous empires, and this period marks the beginning of the formation of the
507 modern Mongolian gene pool. We find that most historic Mongols are well-fitted by a three508 way admixture model with the following ancestry proxies: Ulaanzuuk_SlabGrave, Han, and
509 Alans. Consistent with their PCA location (Fig. 2), Mongol era individuals as a group can be
510 modeled with only 15-18% of western Steppe ancestry (Alan or Sarmatian), but require 55-
511 64% of Ulaanzuuk_SlabGrave and 21-27% of Han-related ancestry (Table S22). Applying the
512 same model to each individual separately, this three-source model adequately explains 56 out
513 of 61 ancient Mongols (based on p-value at threshold of 0.05), as well as one unclassified Late
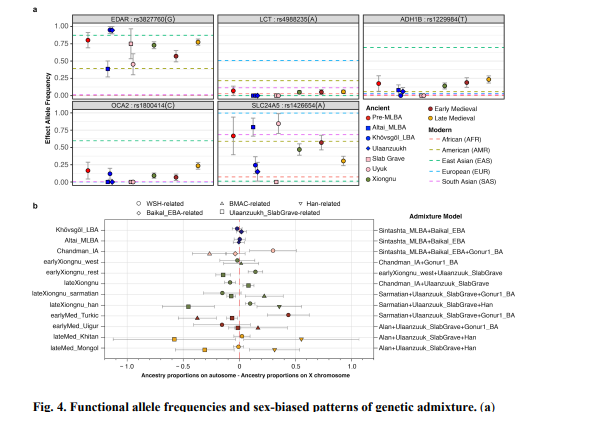
514 Medieval individual dating to around the beginning of the Mongol empire (SHU002) (Table
515 S23).
517 Since the fall of the Mongol empire in 1368 CE, the genetic profile of the Mongolian
518 populations has not substantially changed. The genetic structure established during the Mongol
519 empire continues to characterize present-day Mongolic-speaking populations living in both
520 Mongolia and Russia. We examined the genetic cladality between the historic Mongols and
521 seven present-day Mongolic-speaking groups (Mongols, Kalmyk, Buryat, Khamnegan, Daur,
522 Tu and Mongola) using an individual-based qpWave analysis {Sample; Mongolic group}.
523 Within the resolution of current data, 34/61 historic Mongols are genetically cladal with at least
524 one modern Mongolic-speaking population (Fig. S15). In addition, nearly a third of historic
525 Mongol males (12/38) have Y haplogroup C2b, which is also widespread among modern
526 Mongolians (Table S6, Fig. S3); C2b is the presumed patrilineage of Genghis Khan (Zerjal et
527 al., 2003). The Mongol empire had a profound impact on restructuring the political and genetic
528 landscape of the Eastern Steppe, and these effects endured long after the decline of the empire
529 and are still evident in Mongolia today.
530
531 Functional and gendered aspects of recurrent admixture in the Eastern Steppe
532
533 To investigate the functional aspects of recurrent admixture on the Eastern Steppe, we
534 estimated the population allele frequency of five SNPs associated with functional or
535 evolutionary aspects of lactose digestion (LCT/MCM6), dental morphology (EDAR),
536 pigmentation (OCA2, SLC24A5), and alcohol metabolism (ADH1B) (Fig. 4a). First, we find
537 that despite a pastoralist lifestyle with widespread direct evidence for milk consumption (Jeong
538 et al., 2018; Wilkin et al., 2019), the MLBA and EIA individuals of the Eastern Steppe did not
539 have any derived mutations conferring lactase persistence (LP). Individuals from subsequent
540 periods did have the derived mutation that is today widespread in Europe (rs4988235), but at
541 negligibly low frequency (~5%) and with no increase in frequency over time (Fig. 4a). This is
542 somewhat remarkable given that, in addition to other dairy products, contemporary Mongolian
543 herders consume up to 4-10 liters of airag (fermented mare’s milk, ~2.5% lactose) per day
544 during the summer months (Bat-Oyun et al., 2015), resulting in a daily intake of 100-250g of
545 lactose sugar. Petroglyph depictions of airag production date back to the EIA in the Yenisei
546 Basin (Dėvlet, 1976), and accounts of the historic Mongols record abundant and frequent
547 consumption of airag, as well as a wide range of additional liquid and solid ruminant dairy
548 products (Bayarsaikhan, 2016; Onon, 2005), which has been additionally confirmed by ancient
549 proteomic evidence (Jeong et al., 2018; Wilkin et al., 2019). How Mongolians have been able
550 to digest such large quantities of lactose for millennia in the absence of LP is unknown, but it
551 may be related to their reportedly unusual gut microbiome structure, which today is highly
552 enriched in lactose-digesting Bifidobacteria spp. (Liu et al., 2016).
553
554 Genetic markers that underwent regional selective sweeps show allele frequency changes that
555 correlate with changes in the genome-wide ancestry profile (Fig. 4a). For example, rs3827760
556 in EDAR (ectodysplasin A receptor) and rs1426654 in SLC24A5 (solute carrier family 24
557 member 5) are well-known targets of positive selection in East Asians and Western Eurasians,
558 respectively (Sabeti et al., 2007). Our MLBA and EIA populations show a strong population
559 differentiation in the allele frequencies of these two SNPs: rs3827760 frequency is much higher
560 in groups with higher eastern Eurasian affinity (Khovsgol_LBA, Ulaanzuuk_SlabGrave) while
561 rs1426654 is higher in Altai_MLBA and Chandman_IA (Table S8). We find that two SNPs
562 that have undergone more recent positive selection (Donnelly et al., 2012; Li et al., 2011)
563 among East Asians, rs1229984 in ADH1B (aldehyde dehydrogenase 1B) and rs1800414 in
564 OCA2 (oculocutaneous albinism II), were absent or in extremely low frequency during the
565 MLBA and EIA, when the eastern Eurasian ancestry was primarily ANA-related, but increased
566 in frequency over time as the proportion of East Asian ancestry increased through interactions
567 with China and other groups (Table S8).
568
569 Finally, we investigated gendered dimensions of the population history of the Eastern Steppe.
570 Sex-biased patterns of genetic admixture can be informative about gendered aspects of
571 migration, social kinship, and family structure. We observed an especially strong male-biased
572 admixture from WSH groups during the EIA and Early Medieval periods. To estimate this sex
573 biased admixture, we compared the ancestry proportions in the autosomes and X chromosome
574 for each component, and calculated the statistical significance of their difference following a
575 published strategy (Mathieson et al., 2018). When the proportion of an ancestry is higher in the
576 autosomes than in the X-chromosome, the admixture process involves male ancestors
577 harboring this ancestry in higher proportion than female ancestors. As shown in Fig. 4b, we
578 observe a clear signal of male-biased WSH admixture among the EIA Uyuk and during the
579 Türkic period (i.e., more positive Z scores), which also corresponds to the decline in the Y
580 chromosome lineage Q1a and the concomitant rise of the western Eurasian lineages such as R
581 and J (Fig. S3). During the later Khitan and Mongol empires, sex-bias Z scores shift to indicate
582 a prominant male bias for East Asian-related ancestry (Fig. S6), which can also be seen from
583 the rise in frequency of Y-chromosomal lineage O2a (Fig. S3). The Xiongnu period exhibits
584 the most complex pattern of male-biased admixture, whereby different genetic subsets of the
585 population exhibit evidence of different sources of male-biased admixture (Fig. S6). We also
586 detect ten genetic relative pairs among the Xiongnu individuals in this study, including a father587 daughter pair buried in the same grave (JAG001 and JAA001) at Jargalantyn Am, as well as a
588 mother-son pair (IMA002 and IMA005) at Il'movaya Pad, a brother-sister pair (TMI001 and
589 BUR003) at Tamiryn Ulaan Khoshuu, and a brother-brother pair (SKT002 and SKT006) at
590 Salkhityn Am (Table S7). Of the remaining six pairs, three are female-female relative pairs
591 buried within the same site, suggesting the presence of extended female kinship within
592 Xiongnu groups. These relationships, when combined with mortuary features, offer the first
593 clues to local lineage and kinship structures within the Xiongnu empire, which are otherwise
594 poorly understood.
|
|

