|
|
Post by Admin on Apr 5, 2020 4:14:35 GMT
596 Discussion
597
598 The population history of the Eastern Steppe is one marked by the repeated mixing of diverse
599 eastern and western Eurasian gene pools. However, rather than simple waves of migration,
600 demographic events on the Eastern Steppe have been complex and variable. Generating more
601 than 200 genome-wide ancient datasets, we have presented the first genetic evidence of this
602 dynamic population history, from ca. 4600 BCE through the end of the Mongol empire. We
603 found that the Eastern Steppe was sparsely populated by hunter gatherers of ANA and ANE
604 ancestry during the mid-Holocene, and then shifted to a dairy pastoralist economy during the
605 Bronze Age. Migrating Yamnaya/Afanasievo steppe herders, equipped with carts and domestic
606 livestock (Kovalev and Erdenebaatar, 2009), appear to have first introduced ruminant dairy
607 pastoralism ca. 3000 BCE (Wilkin et al., 2019), but surprisingly had little lasting genetic
608 impact, unlike in Europe (Allentoft et al., 2015; Haak et al., 2015; Mathieson et al., 2015).
609 Subsequent Chemurchek pastoralists in the Altai were confirmed in this study to represent a
610 separate migration of dairy pastoralists with ANE and Iranian-related ancestry, who possibly
611 migrated into the Altai region from the south, likely via Xinjiang and/or mountainous Central
612 Asia due to the concentration of Chemurchek burials in this region (Jia and Betts, 2010). By
613 the MLBA, ruminant dairy pastoralism had been adopted by populations throughout the
614 Eastern Steppe (Wilkin et al., 2019), regardless of ancestry, and this subsistence has continued,
615 with the additions of horse milking in the LBA and camel milking in the Mongol period (Wilkin
616 et al., 2019), to the present day (Bat-Oyun et al., 2015; Kindstedt and Ser-Od, 2019).
617 Puzzlingly, however, there is no evidence of selection for lactase persistence over this 5,000-
618 year history, despite the repeated introduction of this genetic trait by subsequent migrations of
619 groups from the west. This suggests a different trajectory of lactose adaptation in Asia that to
620 date remains unexplained.
621
622 During the MLBA, we observed the formation of a tripartite genetic structure on the Eastern
623 Steppe, characterized by the continuation of pre-Bronze ANA ancestry in the east and a cline
624 of genetic variation between pre-Bronze Age ANA-ANE ancestry in the north and increasing
625 proportions of a new Sintashta-related WSH ancestry in the west. The Sintashta, a western
626 forest steppe culture with genetic links to the European Corded Ware cultures (Mathieson et
627 al., 2015), were masters of bronze metallurgy and chariotry (Anthony, 2010), and the
628 appearance of this ancestry on the Eastern Steppe may be linked to the introduction of new
629 (especially horse-related) technologies. DSKC sites in particular show widespread evidence
630 for horse use in transport, and perhaps even riding (Taylor et al. 2015), and genetic analysis
631 has demonstrated a close link between these animals and the Sintashta chariot horses (Fages et
632 al., 2019). The strong east-west genetic division among Bronze Age Eastern Steppe populations
633 at this time was maintained for more than a millennium and through the end of the EIA, when
634 the first clear evidence for widespread horseback riding appears (Drews, 2004) and the
635 heightened mobility of some groups, notably the eastern Slab Grave culture (Honeychurch,
636 2015), began to disrupt this structure. Eventually, the three major ancestries met and mixed,
637 and this was contemporaneous with the emergence of the Xiongnu empire. The Xiongnu are
638 characterized by extreme levels of genetic heterogeneity and increased diversity as new and
639 additional ancestries from China, Central Asia and the Western Steppe (Sarmatian-related)
640 rapidly entered the gene pool.
641
642 Genetic data for the subsequent early medieval period are relatively sparse and uneven, and
643 few Xianbei or Rouran sites have yet been identified during the 400-year gap between the
644 Xiongnu and Türkic periods. We observed high genetic heterogeneity and diversity during the
645 Türkic and Uyghur periods, with slight shifts in their Sarmatian-like western ancestry
646 component towards that seen among the contemporaneous Alans, a Sarmatian-descendent
647 group known for invading the Roman empire and warring with the Germanic tribes of Europe
648 (Bachrach, 1973). Following the collapse of the Uyghur empire, we documented a final major
649 genetic shift during the late medieval period towards greater eastern Eurasian ancestry, which
650 is consistent with historically documented expansions of Tungusic- (Jurchen) and Mongolic651 (Khitan and Mongol) speaking groups from the northeast into the Eastern Steppe (Biran, 2012).
652 We also observed that this East Asian-related ancestry was brought into the Late Medieval
653 populations more by male than female ancestors, and we observed a corresponding increase in
654 Y-haplogroups associated with southeast Asians and the purported patriline of Ghenghis Khan
655 (O2a and C2b, respectively). By the end of the Mongol period the genetic make-up of the
656 Eastern Steppe had dramatically changed, retaining little of the ANE ancestry that had been a
657 prominent feature during its prehistory. Today, ANE ancestry survives in appreciable amounts
658 only in isolated Siberian groups and among the indigenous peoples of the Americas (Jeong et
659 al., 2019). The genetic profile of the historic Mongols is still reflected among contemporary
660 Mongolians, suggesting a relative stability of this gene pool over the last ~700 years.
661
662 Having documented key periods of genetic shifts in the Eastern steppe, future work may be
663 able to explore whether these shifts are also linked to cultural and technological innovations
664 and how these innovations may have influenced the political landscape. Integrating these
665 findings with research on changes in horse technology and herding practices, as well as shifts
666 in livestock traits and breeds, may prove particularly illuminating. This study represents the
667 first large-scale paleogenomic investigation of the eastern Eurasian Steppe and it sheds light
668 on the remarkably complex and dynamic genetic diversity of the region. Despite this progress,
669 there is still a great need for further genetic research in central and eastern Eurasia, and
670 particularly in northeastern China, the Tarim Basin, and the eastern Kazakh steppe, in order to
671 fully reveal the population history of the Eurasian Steppe and its pivotal role in world
672 prehistory.
|
|
|
|
Post by Admin on Aug 19, 2020 20:34:13 GMT
The Genomic Formation of Human Populations in East Asia
88 The deep population history of East Asia remains poorly understood due to a
89 lack of ancient DNA data and sparse sampling of present-day people. We report
90 genome-wide data from 191 individuals from Mongolia, northern China,
91 Taiwan, the Amur River Basin and Japan dating to 6000 BCE - 1000 CE, many
92 from contexts never previously analyzed with ancient DNA. We also report 383
93 present-day individuals from 46 groups mostly from the Tibetan Plateau and
94 southern China. We document how 6000-3600 BCE people of Mongolia and the
95 Amur River Basin were from populations that expanded over Northeast Asia,
96 likely dispersing the ancestors of Mongolic and Tungusic languages. In a time
97 transect of 89 Mongolians, we reveal how Yamnaya steppe pastoralist spread
98 from the west by 3300-2900 BCE in association with the Afanasievo culture,
99 although we also document a boy buried in an Afanasievo barrow with ancestry
100 entirely from local Mongolian hunter-gatherers, representing a unique case of
101 someone of entirely non-Yamnaya ancestry interred in this way. The second
102 spread of Yamnaya-derived ancestry came via groups that harbored about a
103 third of their ancestry from European farmers, which nearly completely
104 displaced unmixed Yamnaya-related lineages in Mongolia in the second
105 millennium BCE, but did not replace Afanasievo lineages in western China
106 where Afanasievo ancestry persisted, plausibly acting as the source of the early
107 splitting Tocharian branch of Indo-European languages. Analyzing 20 Yellow
108 River Basin farmers dating to ~3000 BCE, we document a population that was a
109 plausible vector for the spread of Sino-Tibetan languages both to the Tibetan
110 Plateau and to the central plain where they mixed with southern agriculturalists
111 to form the ancestors of Han Chinese. We show that the individuals in a time
112 transect of 52 ancient Taiwan individuals spanning at least 1400 BCE to 600 CE
113 were consistent with being nearly direct descendants of Yangtze Valley first
114 farmers who likely spread Austronesian, Tai-Kadai and Austroasiatic languages
115 across Southeast and South Asia and mixing with the people they encountered,
116 contributing to a four-fold reduction of genetic differentiation during the
117 emergence of complex societies. We finally report data from Jomon hunter
118 gatherers from Japan who harbored one of the earliest splitting branches of East
119 Eurasian variation, and show an affinity among Jomon, Amur River Basin,
120 ancient Taiwan, and Austronesian-speakers, as expected for ancestry if they all
121 had contributions from a Late Pleistocene coastal route migration to East Asia.
122
123 Main text
124 East Asia, one of the oldest centers of animal and plant domestication, today harbors
125 more than a fifth of the world’s human population, with present-day groups speaking
126 languages representing eleven major families: Sino-Tibetan, Tai-Kadai, Austronesian,
127 Austroasiatic, Hmong-Mien, Indo-European, Altaic (Mongolic, Turkic, and
128 Tungusic), Koreanic, Japonic, Yukgahiric, and Chukotko-o-Kanchatkan1. The past
129 10,000 years have been a period of profound economic and cultural change in East
130 Asia, but our current understanding of the genetic diversity, major mixture events, and
131 population movements and turnovers during the transition from foraging to
132 agriculture remains poor due to minimal sampling of the diversity of present-day
133 people on the Tibetan Plateau and southern China. A particular limitation has been a
134 deficiency in ancient DNA data, which has been a powerful tool for discerning the
135 deep history of populations in Western and Central Eurasia3-8.
137 We genotyped 383 present-day individuals from 46 populations indigenous to China
138 (n=337) and Nepal (n=46) using the Affymetrix Human Origins array (Table S1 and
139 Supplementary Information section 1). We also report genome-wide data from 191
140 ancient East Asians, many from cultural contexts for which there is no published
141 ancient DNA data. From Mongolia we report 89 individuals from 52 sites dating
142 between ~6000 BCE to ~1000 CE. From China we report 20 individuals from the
143 ~3000 BCE Neolithic site of Wuzhuangguoliang. From Japan we report 7 Jomon
144 hunter-gatherers from 3500-1500 BCE. From the Russian Far East we report 23
145 individuals: 18 from the Neolithic Boisman-2 cemetery at ~5000 BCE, 1 from the
146 Iron Age Yankovsky culture at ~1000 BCE, 3 from the Medieval Heishui Mohe and
147 Bohai Mohe culture at ~1000 CE; and 1 historic period hunter-gatherer from Sakhalin
148 Island. From archaeological sites in Eastern Taiwan—the Bilhun site at Hanben on the
149 main island and the Gongguan site on Green Island—we report 52 individuals from
150 the Late Neolithic through the Iron Age spanning at least 1400 BCE - 600 CE.
|
|
|
|
Post by Admin on Aug 20, 2020 7:03:57 GMT

152 For all but the Chinese samples we enriched the ancient DNA for a targeted set of
153 about 1.2 million single nucleotide polymorphisms (SNPs)4,9, while for the
154 Wuzhuangguoliang samples from China we used exome capture (18 individuals) or
155 shotgun sequencing (2 individuals) (Figure 1, Supplementary Data files 1 and 2 and
156 Supplementary Information section 1). We performed quality control to test for
157 contamination by other human sequences, assessed by the rate of cytosine to thymine
158 substitution in the terminal nucleotide and polymorphism in mitochondrial DNA
159 as well as X chromosome sequences in males, and restricted analysis to
160 individuals with minimal contamination11(Online Table 1). We detected close kinship
161 between individuals at the same site, including a Boisman nuclear family with 2
162 parents and 4 children (Table S2). We merged the new data with previously reported
163 data: 4 Jomon individuals, 8 Amur River Basin Neolithic individuals from the Devil’s
164 Gate site, 72 individuals from the Neolithic to the Iron Age in Southeast Asia, and 8
165 from Nepal7,12-20. We assembled 123 radiocarbon dates using bone from the
166 individuals, of which 94 are newly reported (Online Table 3), and clustered
167 individuals based on time period and cultural associations, then further by genetic
168 cluster which in the Mongolian samples we designated by number (our group names
169 thus have the format “<Country>_<Time Period>_<Genetic Cluster>_<Cultural
170 Association If Any>”) (Supplementary Note, Table S1 and Online Table 1). We
171 merged the data with previously reported data (Online Table 4).
|
|
|
|
Post by Admin on Aug 20, 2020 21:13:59 GMT
173 We carried out Principal Component Analysis (PCA) using smartpca21, projecting the
174 ancient samples onto axes computed using present-day people. The analysis shows
175 that population structure in East Asia is correlated with geographic and linguistic
176 categories, albeit with important exceptions. Groups in Northwest China, Nepal, and
177 Siberia deviate towards West Eurasians in the PCA (Supplementary Information
178 section 2, Figure 2), reflecting multiple episodes of West Eurasian-related admixture
179 that we estimate occurred 5 to 70 generations ago based on the decay of linkage
180 disequilibrium22(Table S3 and Table S4). East Asians with minimal proportions of
181 West Eurasian-related ancestry fall along a gradient with three clusters at their poles.
182 The “Amur Basin Cluster” correlates geographically with ancient and present-day
183 populations living in the Amur River Basin, and linguistically with present-day
184 indigenous people speaking Tungusic languages and the Nivkh. The “Tibetan Plateau
185 Cluster” is most strongly represented in ancient Chokhopani, Mebrak, and Samzdong
186 individuals from Nepal15 and in present-day people speaking Tibetan-Burman
187 languages and living on the Tibetan Plateau. The “Southeast Asian Cluster” is
188 maximized in ancient Taiwan groups and present-day people in Southeast Asia and
189 southern parts of China speaking Austroasiatic, Tai-Kadai and Austronesian
190 languages (Figure S1, Figure S2). Han are intermediate among these clusters, with
191 northern Han projecting close to the Neolithic Wuzhuangguoliang individuals from
192 northern China (Figure 2). We observe two genetic clusters within Mongolia: one falls
193 closer to ancient individuals from the Amur Basin Cluster (‘East’ based on their
194 geography), and the second clusters toward ancient individuals of the Afanasievo
195 culture ( ‘West’), while a few individuals take intermediate positions between the two
196 (Supplementary Information section 2).
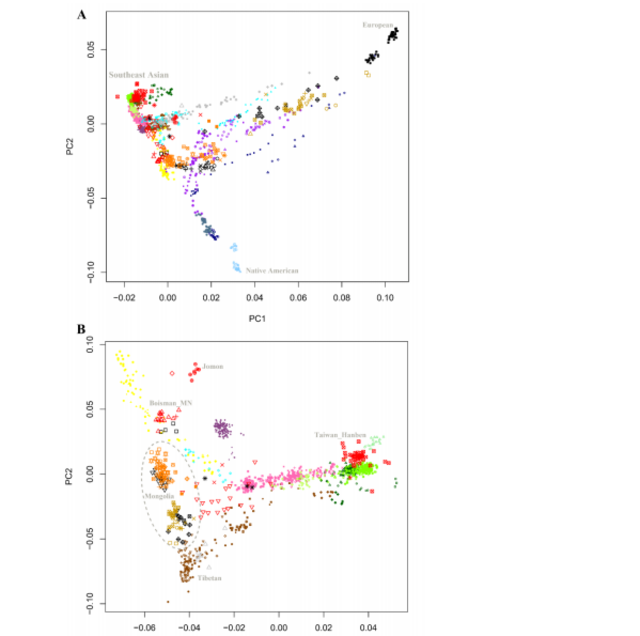 Figure 2: Principal Component Analysis (PCA). (A) Projection of ancient samples onto PCA dimensions 1 and 2 defined by East Asians, Europeans, Siberians and Native Americans. (B) Projection onto groups with the little West Eurasian mixture.
198 The three most ancient individuals of the Mongolia ‘East’ cluster are from the
199 Kherlen River region of eastern Mongolia (Tamsag-Bulag culture) and date to 6000-
200 4300 BCE (this places them in the Early Neolithic period, which in Northeast Asia is
201 defined by the use of pottery and not by agriculture23). These individuals are
202 genetically similar to previously reported Neolithic individuals from the cis-Baikal
203 region and have minimal evidence of West Eurasian-related admixture as shown in
204 PCA (Figure 2), f4-statistics and qpAdm (Table S5, Online Table 5, labeled as
205 Mongolia_East_N). The other seven Neolithic hunter-gatherers from northern
206 Mongolia (labeled as Mongolia_North_N) can be modeled as having 5.4% ±1.1%
207 ancestry from a source related to previously reported West Siberian Hunter-gatherers
208 (WSHG)8(Online Table 5), consistent with the PCA where they are part of an east
209 west Neolithic admixture cline in Eurasia with increasing proximity to West Eurasians
210 in groups further west. Because of this ancestry complexity, we use the
211 Mongolia_East_N individuals without significant evidence of West Eurasian-related
212 admixture as reference points for modeling the East Asian-related ancestry in later
213 groups (Online Table 5). The two oldest individuals from the Mongolia ‘West’ cluster
214 have very different ancestry: they are from the Shatar Chuluu kurgan site associated
215 with the Afanasievo culture, with one directly dated to 3316-2918 calBCE (we quote
216 a 95% confidence interval here and in what follows whenever we mention a direct
217 date), and are indistinguishable in ancestry from previously published ancient
218 Afanasievo individuals from the Altai region of present-day Russia, who in turn are
219 similar to previously reported Yamnaya culture individuals supporting findings that
220 eastward Yamnaya migration had a major impact on people of the Afansievo culture.
221 All the later Mongolian individuals in our time transect were mixtures of
222 Mongolian Neolithic groups and more western steppe-related sources, as reflected by
223 statistics of the form f3 (X, Y; Later Mongolian Groups), which resulted in
224 significantly negative Z scores (Z<−3) when Mongolia_East_N was used as X, and
225 when Yamnaya-related Steppe populations, AfontovaGora3, WSHG, or European
226 Middle/Late Neolithic or Bronze Age populations were used as Y (Table S6).
|
|
|
|
Post by Admin on Aug 21, 2020 20:25:17 GMT
228 To quantify the admixture history of the later Mongolians, we again used qpAdm. A
229 large number of groups could be modeled as simple two-way admixtures of
230 Mongolia_East_N as one source (in proportions of 65-100%) and WSHG as the other
231 source (in proportions of 0-35%), with negligible contribution from Yamnaya-related
232 sources as confirmed by including Russia_Afanasievo and Russia_Sintashta groups in
233 the outgroup set (Figure 3). The groups that fit this model were not only the two
234 Neolithic groups (0-5% WSHG), but also the Early Bronze Age people from the
235 Afanasievo Kurgak govi site (15%), the Ulgii group (28%), the main grouping of
236 individuals from the Middle Bronze Age Munkhkhairkhan culture (33%), Late Bronze
237 Age burials of the Ulaanzuukh type (6%), a combined group from the Center-West
238 region (27%), the Mongun Taiga type from Khukh tolgoi (35%), and people of the
239 Iron Age Slab Grave culture (9%). A striking finding in light of previous
240 archaeological and genetic data is that the male child from Kurgak govi (individual
241 I13957, skeletal code AT_629) has no evidence of Yamnaya-related ancestry despite
242 his association with Afanasievo material culture (for example, he was buried in a
243 barrow in the form of circular platform edged by vertical stone slabs, in stretched
244 position on the back on the bottom of deep rectangular pit and with a typical
245 Afanasievo egg-shaped vessel (Supplementary Note); his late Afanasievo chronology
246 is confirmed by a direct radiocarbon date of 2858-2505 BCE24). This is the first
247 known case of an individual buried with Afanasievo cultural traditions who is not
248 overwhelmingly Yamnaya-related, and he also shows genetic continuity with an
249 individual buried at the same site Kurgak govi 2 in a square barrow (individual I6361,
250 skeletal code AT_635, direct radiocarbon date 2618-2487 BCE). We label this second
251 individuals as having an Ulgii cultural association, although a different archaeological
252 assessment associates this individual to the Afanasievo or Chemurchek cultures25, so
253 it is possible that this provides a second example of Afanasievo material culture being
254 adopted by individuals without any Yamnaya ancestry. The legacy of the Yamnaya
255 era spread into Mongolia continued in two individuals from the Chemurchek culture
256 whose ancestry can be only modeled by using Afanasievo as one of the sources
257 (49.0%±2.6%, Online Table 5). This model fits even when ancient European farmers
258 are included in the outgroups, showing that if the long-distance transfer of West
259 European megalithic cultural traditions to people of the Chemurchek culture that has
260 been suggested in the archaeological literature occurred, it must have been through
261 spread of ideas rather than through movement of people

263 Beginning in the Middle Bronze Age in Mongolia, there is no compelling evidence
264 for a persistence of the Yamnaya-derived lineages originally spread into the region
265 with Afanasievo. Instead in the Late Bronze Age and Iron Age and afterward we have
266 data from multiple Mongolian groups whose Yamnaya-related ancestry can only be
267 modeled as deriving not from the initial Afanasievo migration but instead from a later
268 eastward spread into Mongolia related to people of the Middle to Late Bronze Age
269 Sintashta and Andronovo horizons who were themselves a mixture of ~2/3 Yamnayarelated
270 and 1/3 European farmer-related,7,8. The Sintashta-related ancestry is
271 detected in proportions of 5% to 57% in individuals from the
272 Mongolia_LBA_6_Khovsgol (a culturally mixed group from the literature14),
273 Mongolia_LBA_3_MongunTaiga, Mongolia_LBA_5_CenterWest,
274 Mongolia_EIA_4_Sagly, Mongolia_EIA_6_Pazyryk, and Mongolia_Mongol groups,
275 with the most substantial proportions of Sintashta-related ancestry always coming
276 from western Mongolia (Figure 3, Online Table 5). For all these groups, the qpAdm
277 ancestry models pass when Afanasievo is included in the outgroups while models
278 with Afanasievo treated as the source with Sintashta more distantly related outgroups
279 are all rejected (Figure 3, Online Table 5). Starting from the Early Iron Age, we
280 finally detect evidence of gene flow in Mongolia from groups related to Han Chinese.
281 Specifically, when Han are included in the outgroups, our models of mixtures in
282 different proportions of Mongolia_East_N, Russia_Afanasievo, Russia_Sintashta, and
283 WSHG continue to work for all Bronze Age and Neolithic groups, but fail for an
284 Early Iron Age individual from Tsengel sum (Mongolia_EIA_5), and for Xiongnu and
285 Mongols. When we include Han Chinese as a possible source, we estimate ancestry
286 proportions of 20-40% in Xiongnu and Mongols (Online Table 5).
|
|

